概論
細胞老化の新局面〜より完全な理解と健康寿命延伸に向けた可能性
New era of cellular senescence: a deeper understanding and its potential towards extension of healthy lifespan
原 英二
Eiji Hara:Department of Molecular Microbiology, Research Institute for Microbial Diseases(RIMD),Osaka University/Laboratory of Aging Biology, Immunology Frontier Research Center(IFReC),Osaka University(大阪大学微生物病研究所遺伝子生物学分野/大阪大学免疫学フロンティア研究センター老化生物学)
われわれの身体を構成する細胞は,異常を感知すると細胞分裂を停止する安全装置を備えている.細胞老化はこの安全装置の一つであり,長い間がん化の危険がある異常細胞の増殖を防ぐがん抑制機構として働いていると考えられてきた.2003年に本誌で細胞老化の特集を企画した際には細胞老化のがん抑制機構としての作用を中心に概説した.しかし,あれから16年,細胞老化研究は大きく発展し,単に細胞分裂を停止する現象としてだけではなく,さまざまな生理活性物質を分泌するSASPという現象を起こすことが明らかになってきた.SASPを介して個体発生,組織修復,免疫制御など生体の恒常性維持に寄与する反面,過度なSASPは慢性炎症を惹起して生体機能の低下やがんなどの加齢性疾患の発症を促進する副作用があることも明らかになってきた.本特集では,細胞老化のより完全な理解とその創薬応用に向けた可能性について概説する.
はじめに
哺乳動物の正常な体細胞を継代培養すると,細胞分裂をある一定の回数くり返した後,不可逆的に細胞分裂を停止することが1956年にHaffとSwimらによって報告された1).この報告はほとんど注目されることはなかったが,その5年後にHayflickらによりヒトの体細胞(線維芽細胞)を用いて同様の実験結果が報告され,この現象に“cellular senescence(細胞老化)”という名前が付けられると多くの研究者の注目を集めるようになった2).細胞老化は細胞の分裂回数を制限することでがん化を防いでいる可能性や,個体老化の原因となっている可能性が指摘される一方で,正常な細胞を生体からとり出して培養したことにより生じるアーティファクトではないかという懐疑的な意見も多く,細胞老化の生理作用については長い間結論がでないままであった.しかし,その後,細胞老化誘導に関わる分子機構が明らかになるにつれ,生体内でも細胞老化と類似の現象が起こっている可能性が報告され,細胞老化の生体内での役割が注目されるようになった3).さらに,最近の研究により細胞老化を起こした細胞(以下,老化細胞とよぶ)は単に細胞分裂を停止しているだけではなく,炎症性サイトカイン,ケモカイン,細胞外マトリクス分解酵素や増殖因子,さらには分泌膜小胞(エクソソーム)等さまざまな生理活性物質を分泌するsenescence-associated secretary phenotype(SASP)とよばれる現象を起こしていることも明らかになり4),生体の恒常性維持に寄与しているだけでなく状況によっては恒常性を破綻させる副作用を示すことが明らかになってきた5).事実,遺伝子改変マウスを用いて加齢の過程で体内に蓄積した老化細胞を死滅させると,がんを含めたさまざまな加齢性疾患の発症率が低下し,健康寿命が延伸するという報告がある6).また,肥満マウスにおいて肝臓に生じた老化細胞を除去すると肝がんの発症が低下することも示されており7),そのヒトへの応用が期待されている.
近年,先進国においては平均寿命の延長に伴い,がん,認知症,心・血管系疾患などのさまざまな老化関連疾患が増え,高齢者のQOLの低下のみならず,医療費や介護費の増加などが大きな社会問題になりつつある.特に少子高齢化が著しい日本においてはこの問題は深刻であり,効果的な対策が求められている.このような社会背景と細胞老化研究の進展状況をかんがみ,本特集ではいま,大きく広がりつつある細胞老化の生理作用の解明とその応用に向けた可能性に焦点を当て,この分野において第一線で活躍しておられる研究者に執筆を依頼した.また,比較生物学的な観点から細胞老化を検証することで細胞老化のより深い理解とその老化関連疾患制御に向けた可能性の発見に資することができれば幸甚の至りである.
1細胞老化誘導メカニズムとその多様性
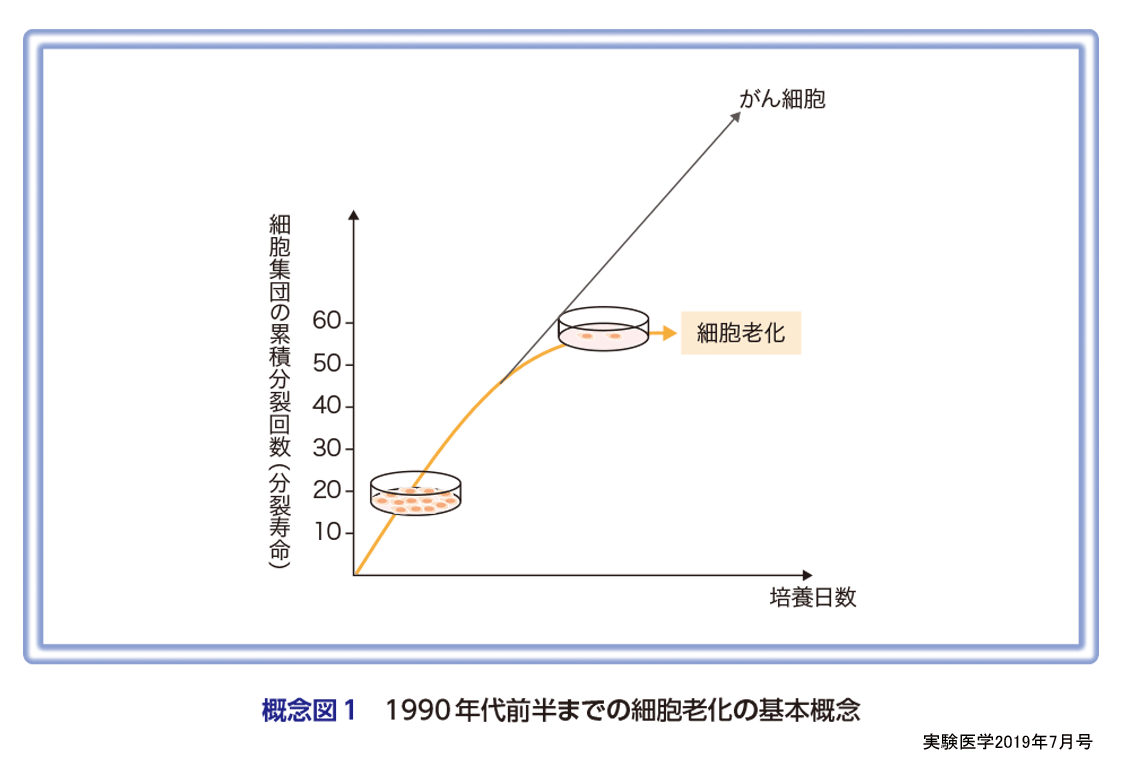
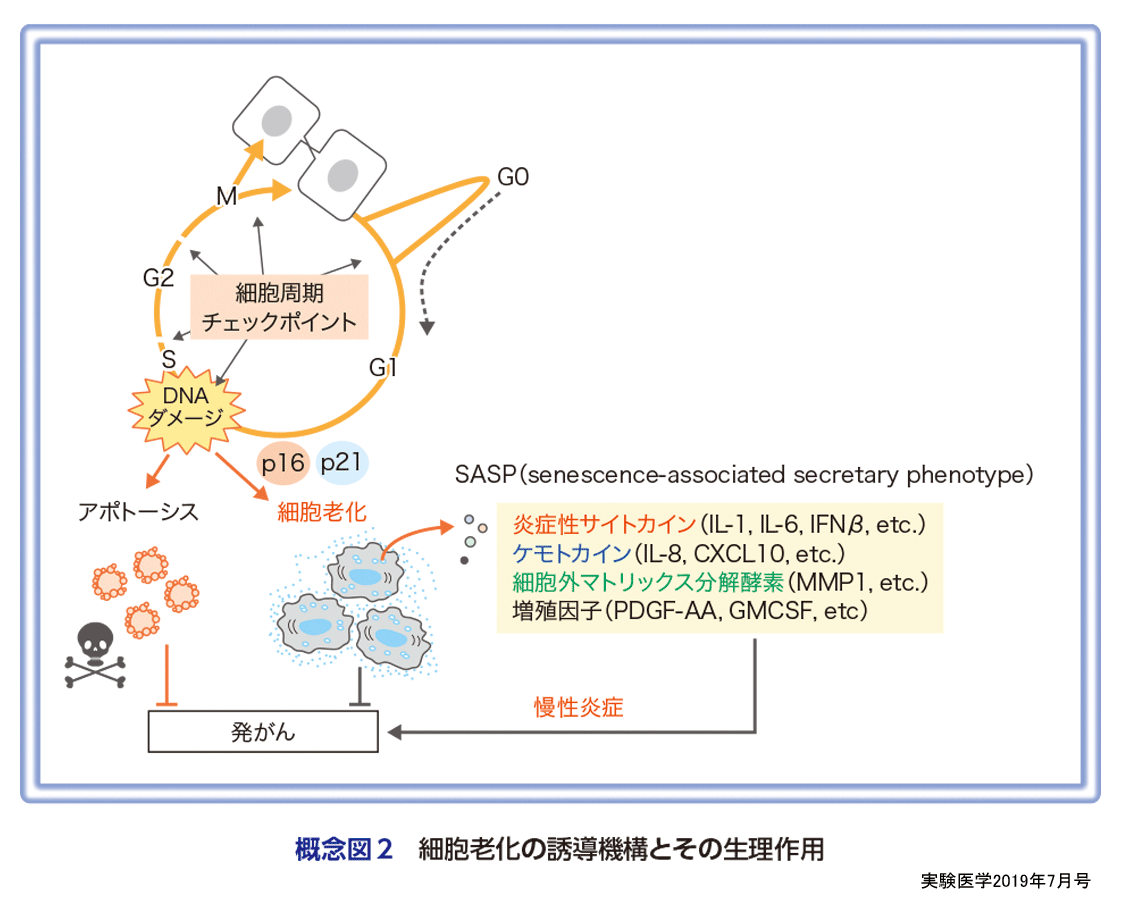
細胞老化はもともと正常なヒトの体細胞が継代培養をくり返すことで分裂増殖能を喪失する現象として定義された(概念図1)2).このため,正常な体細胞には分裂回数をカウントするシステムと細胞分裂を停止させる実行因子が備わっていると考えられ,その実体解明に向けた研究が行われてきた.1990年代になり,細胞の分裂回数カウンティングシステムとしてテロメア長の短小化が8),そして細胞分裂を停止させる実行因子としてp16INK4aやp21Waf1/Cip1/SdI1などのサイクリン依存性キナーゼ阻害因子(CDKIs)が重要な役割を果たしていることが明らかになってきた9)10).一方,マウスの体細胞ではヒトの体細胞と同様に継代培養により細胞老化様の増殖停止を起こすが,テロメア長の短小化は起こらない.また,低酸素条件下で培養すると細胞老化が起こらなくなることから,マウスの細胞は主に酸化的ストレスによって細胞老化様の増殖停止を起こすと考えられる11).じつは,ヒトの線維芽細胞も古くから培養中の酸素濃度を低くすると分裂可能な回数が延長することが報告されており12),ヒトの細胞でもテロメアの短小化だけが細胞老化の誘導時期を決定しているわけではない.また,1990年代後半になるとp16INK4aやp21Waf1/Cip1/SdI1など細胞老化の誘導にかかわる因子の解析から,正常な細胞に発がんの危険性があるストレス(がん遺伝子の活性化や過度な酸化的ストレス,放射線・紫外線照射等)が生じると,継代培養をくり返さなくても,すみやかに細胞老化様の増殖停止が起こることが報告されるようになってきた13).さらに,継代培養による細胞老化に加え,これら発がんストレスにより急速に起こる細胞老化様の増殖停止に共通してDNA損傷応答が重要な役割を果たしていることが明らかになってきた(概念図2)14).また,ストレスの種類や細胞の種類,さらには細胞環境によって,細胞分裂を停止させる実行因子が多少異なることも報告されている.例えば,p16INK4aやp21Waf1/Cip1/SdI1の代わりにそれらのファミリー分子が働いたり,サイクリンやサイクリン依存性キナーゼ(CDK)の発現自体が減少することで細胞周期の進行が停止する場合があることが報告されている15).また,老化細胞では増殖停止を起こしてからの時間に応じて染色体のエピジェネティックな修飾状態が異なり,発現する遺伝子も変化するため,細胞分裂の停止状態を維持する機構も異なってくることが報告されている16)〜19)(城村らの稿).すなわち,老化細胞と一言で言っても細胞種やストレスによってかなり異なる状態の細胞であり,生体内においてはじつに多様な老化細胞が存在していることが予想される.
2細胞老化の生理作用
正常細胞と異なり,がん細胞の多くが細胞老化を起こさず無限に継代培養可能であることから,細胞老化の生理作用として発がん抑制の側面が古くから注目されてきた3).p16INK4a,サイクリンDキナーゼ,RB,p53等,細胞老化の誘導にかかわる中心的な遺伝子の多くがヒトのがん細胞において高頻度に遺伝子異常を起こしていることからも細胞老化の重要な生理作用の一つとしてがん抑制機構があげられる(太田・山田の稿).興味深いことにp16INK4a遺伝子を欠失したノックアウトマウスは若いときには異常がみられないが,1年齢を超えたあたりから発がんの頻度が増え野生型マウスに比べ早く死亡する20).このことからも,細胞老化が加齢に伴う発がんを抑制していることが窺える.一方,p16INK4a遺伝子の発現は加齢とともにマウスのさまざまな臓器で上昇することが知られているが,21)22),ノックアウトマウスは血液,膵臓,神経などにおいて幹細胞や前駆細胞が増加し,それら組織の再生能力が高まることが報告されている23)〜25).このため,細胞老化はがん抑制機構として働く一方で組織の再生能力を低下させ,個体老化を促進する働きがあると推察されているが,p16INK4aノックアウトマウスが早期にがんを発症し死亡するため20),その証明は未完のままである.
一方,先述のように老化細胞は単に増殖を停止しているだけではなくSASPを起こすことで周囲の細胞にも影響を与える.老化細胞が炎症性サイトカインなどの分泌因子を高発現すること自体は古くから知られていたが26),当時はほとんど注目されなかった.しかし,この現象がSASPと名付けられて以降,その重要性が認識され,いまさらなる広がりを見せている.SASPの生理作用は,どのSASP因子が,どの細胞に作用するかによって大きく異なるが,SASP因子の主要な成分として炎症性サイトカインや増殖因子が含まれることから,働きすぎると生体にとってよい作用だけでなく,慢性炎症や発がんを引き起こすことが知られておりその制御が重要である4)5)(高橋・原の稿).また,遺伝子改変によりp16INK4a遺伝子を発現する細胞を死滅させることができるINK-ATTACマウスを用いた解析により,p16INK4a発現細胞を除去することで老化細胞が体内に蓄積しないようにしておくと,動脈硬化,白内障,サルコペニアや発がんなどさまざまな加齢性疾患の発症率が低下し,健康寿命が延伸することが報告され6),加齢性疾患の新たな治療戦略の可能性を秘めていると大きなインパクトを与えた.特にp16陽性老化細胞の除去が動脈硬化の進展を抑制するのみならず退縮させることが示され,心血管系疾患の治療への応用が期待される(勝海・南野の稿).しかし,p16INK4aを発現しているすべての細胞が老化細胞とは限らないことも指摘されており27)28),より詳細な解析と慎重な解釈が必要である(杉本・三河の稿).
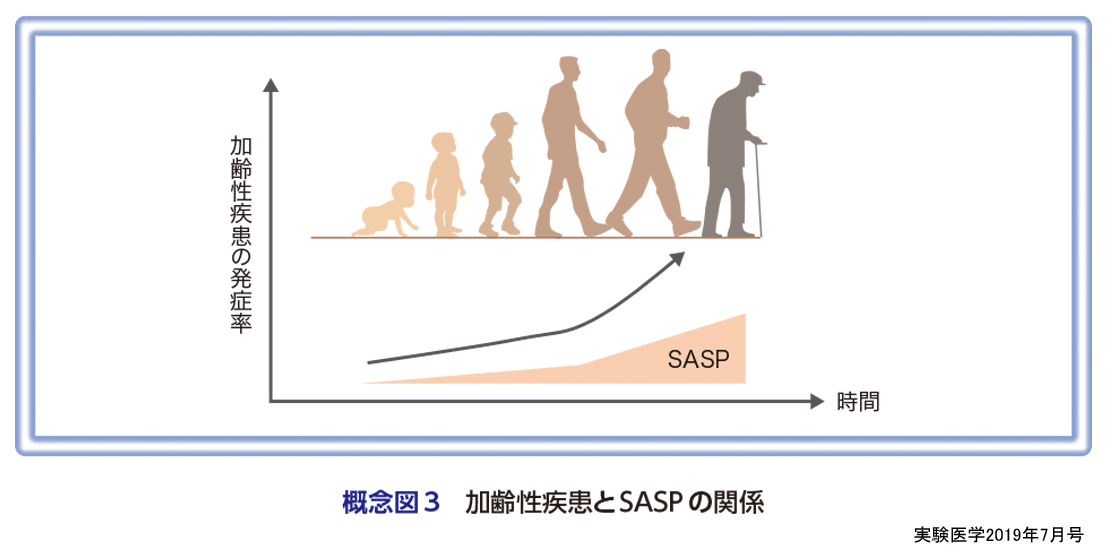
一方,健康長寿ネズミとして知られるハダカデバネズミの細胞は細胞老化を起こすと直ぐにアポトーシスを起こすことが報告されている(河村らの稿)29).また,最近,ショウジョウバエでも細胞老化とSASPが起こり,SASPを介して発がんが促進されることが報告された30)(井藤・井垣の稿).われわれは肥満マウスにおいて肝臓に生じた老化細胞を除去すると肝がんの発症が低下することを報告している7).このように異なるモデル生物や異なるストレス環境下における細胞老化の役割を比較検討すると,加齢に伴う老化細胞の蓄積はSASPを介して生体機能の低下や加齢性疾患の発症を促進している可能性が高い(概念図3).
3細胞老化を標的とした創薬の可能性
細胞老化を標的した創薬の方向性は以下の3つに大別される.
❶ 細胞老化誘導によるがん治療薬の開発
がん細胞ではp16INK4a遺伝子が失活したり,その下流因子であるCDK4/6キナーゼが過度に活性化する変異が多く見つかっており,がん細胞においてCDK4/6の機能を阻害することで細胞老化を誘導してがん治療をめざす抗がん剤の開発が古くから行われてきた.すでにCDK4/6キナーゼ活性を阻害する低分子阻害剤(パルボシクリブやアベマシクリブ)が開発されており,乳がんの治療薬等として使用されている31).
❷ 体内から老化細胞を除去する薬剤(senolytic薬)の開発
INK-ATTACマウスによる研究結果をもとに,健康寿命の延伸をめざして危険な老化細胞を体内から除去するsenolytic(senescenceとlyticの造語)薬の開発が行われている.多種チロシンキナーゼの阻害剤であるダサチニブとフラボノイド系ポリフェノールであるケルセチンの組合わせにsenolytic効果があることが報告されており,マウスにおいては加齢に伴う筋力の衰えや寿命延長効果が報告されている.また,老化細胞では抗アポトーシス作用を示すBclファミリーの発現が亢進していることからBcl阻害剤であるABT263にsenolytic作用があることが報告されている.すでに肺小細胞がんやプラチナ製剤抵抗性卵巣がん等に対する臨床試験が行われているが,血小板を減少させる副作用がみられており,安全性に問題が生じている.このほかにも,老化細胞で亢進している解糖系を標的とする2-deoxy-D-glucose(2-DG)や細胞死抑制作用を有するFOXO4を阻害するペプチド,熱ショックタンパク質(HSP90)の阻害剤アルベスピマイシンなど,多数のsenolytic薬の開発が進んでいる(三河・杉本の稿)32).
❸ SASP誘導阻害(調節)薬の開発
SASP自体は生体にとって有益な作用もあるが,過度に働くことで慢性炎症などの副作用を起こす4)5).このため,SASPの誘導を阻害する薬剤の探索が行われている.すでに抗がん剤として臨床試験が進んでいるBETインヒビターにSASPを抑制する活性があることや33),SASPの誘導に重要な働きをしているcGAS-STING経路の阻害剤などが注目されている.特に既存薬であるアスピリンにcGAS阻害活性が見つかったことは注目に値する34).
おわりに
細胞老化の概念はこの20年の間に大きく変化した.分裂寿命という概念から,さまざまなストレスによって誘導されるストレス応答という概念への変化.また,ストレスの種類や細胞の種類によって引き起こされる反応もいくぶん異なり,老化細胞に多様性があることも明らかになってきた35).そして長い間,細胞老化の主な機能と考えられてきた不可逆的な細胞分裂停止以上に生体内ではSASPが重要な役割を果たしていることも明らかにありつつある.前述のINK-ATTACマウスやsenolytic薬を用いた研究からSASPは生体にとって有益な作用より有害な作用の方が多いように思える.では,なぜ,われわれは進化の過程でSASPを備えたのだろうか? SASP誘導には自然免疫応答(cGAS-STING経路)が利用されていることを考慮すると(高橋・原の稿),やはりSASPは若いうちは生体防御機構として働いているのではないかと想像される.120年前まで先進国の平均寿命が今の約半分であったことを考えると36),おそらく人類はつい最近までSASPの副作用が出る前に死亡していた可能性が高い.すなわち,SASPの副作用に直面する現代社会の状況は進化のスピートをはるかに越えて寿命が伸びたために起こった想定外の状況であると言える.今後人類はAIをはじめ生物学的な進化の法則から外れた数々の想定外と向き合っていかなければいけないことが予想される.細胞老化の副作用もその一つと考えられ,今後さらなる解明が必要である.
文献
- Haff RF & Swim HE:Proc Soc Exp Biol Med, 93:200-204, 1956
- Hayflick L & Moorhead PS:Exp Cell Res, 25:585-621, 1961
- Herbig U & Sedivy JM:Mech Ageing Dev, 127:16-24, 2006
- Rodier F & Campisi J:J Cell Biol, 192:547-556, 2011
- Watanabe S, et al:Cancer Sci, 108:563-569, 2017
- Baker DJ, et al:Nature, 530:184-189, 2016
- Yoshimoto S, et al:Nature, 499:97-101, 2013
- Ishikawa F:Cancer Sci, 104:790-794, 2013
- Noda A, et al:Exp Cell Res, 211:90-98, 1994
- Hara E, et al:Mol Cell Biol, 16:859-867, 1996
- Parrinello S, et al:Nat Cell Biol, 5:741-747, 2003
- Packer L & Fuehr K:Nature, 267:423-425, 1977
- Serrano M & Blasco MA:Curr Opin Cell Biol, 13:748-753, 2001
- Campisi J & d’Adda di Fagagna F:Nat Rev Mol Cell Biol, 8:729-740, 2007
- Ohtani N, et al:Cancer Sci, 100:792-797, 2009
- Takahashi A, et al:Mol Cell, 45:123-131, 2012
- Takahashi A, et al:Nat Cell Biol, 8:1291-1297, 2006
- De Cecco M, et al:Nature, 566:73-78, 2019
- Johmura Y, et al:Mol Cell, 55:73-84, 2014
- Sato S, et al:Nat Commun, 6:7035, 2015
- Krishnamurthy J, et al:J Clin Invest, 114:1299-1307, 2004
- Yamakoshi K, et al:J Cell Biol, 186:393-407, 2009
- Janzen V, et al:Nature, 443:421-426, 2006
- Krishnamurthy J, et al:Nature, 443:453-457, 2006
- Molofsky AV, et al:Nature, 443:448-452, 2006
- 原 英二:細胞工学, 34:1122-1124, 2015
- Hall BM, et al:Aging (Albany NY), 9:1867-1884, 2017
- Okuma A, et al:Nat Commun, 8:2050, 2017
- Miyawaki S, et al:Nat Commun, 7:11471, 2016
- Nakamura M, et al:Nat Commun, 5:5264, 2014
- Schettini F, et al:Front Oncol, 8:608, 2018
- 渡邉すぎ子,原 英二:医学のあゆみ, 267:1143-1149, 2018
- Tasdemir N, et al:Cancer Discov, 6:612-629, 2016
- Dai J, et al:Cell, 176:1447-1460.e14, 2019
- Chan ASL & Narita M:Genes Dev, 33:127-143, 2019
- Kinsella KG:Am J Clin Nutr, 55:1196S-1202S, 1992
著者プロフィール
原 英二:大学院生の頃より細胞老化の研究を開始し,(英国)Imperial Cancer Research Fund LaboratoriesのGordon Peters博士のラボでp16INK4a遺伝子が細胞老化の誘導に関与していることを発見した.1998年に(英国)Cancer Research UK Paterson Instituteでラボヘッドとして独立.その後,徳島大学,がん研究会,を経て2015年より現職.’17年よりAMED老化メカニズムの解明・制御プロジェクト「老化機構・制御研究拠点」拠点長.生体内で細胞老化を誘導するストレスの実体解明とその制御をめざしている.’20年11月2~5日に大阪で開催予定のInternational Cell Senescence Association(ICSA)2020 Conferenceの開催責任者.世界中から細胞老化研究の第一人者が集結しますので,奮ってご参加ください(https://www.cellsenescence.info/).