
概論
相分離研究による細胞像の再構築
廣瀬哲郎
(大阪大学大学院生命機能研究科)
[略語]
- ALS:
- amyotrophic lateral sclerosis(筋萎縮性側索硬化症)
- LLPS:
- liquid-liquid phase separation(液 - 液相分離)
はじめに
20世紀初頭,中枢神経構造の顕微鏡観察によってニューロン説を提唱し,今日の神経科学の基礎を築き上げた偉大な神経解剖学者サンティアゴ・ラモン・イ カハールは,細胞核内の核小体に付随した顆粒状構造体を発見した1).今日では,この構造体はその名を冠してカハール体とよばれている.一方,カハールと主張を異にしたまま同時にノーベル賞を受賞したカミッロ・ゴルジは,細胞内小器官ゴルジ体にその名を残すことになった.カハール体とゴルジ体,大きさも形も局在場所も異なる2つの細胞内小器官(オルガネラ)であるが,決定的な違いはゴルジ体が脂質二重膜によって囲まれているのに対して,カハール体は膜構造をもたない非膜オルガネラである点である.一般的にオルガネラというとゴルジ体に加えてミトコンドリアや葉緑体など膜に囲まれた細胞内小器官を思い浮かべるが,近年,非膜オルガネラが大きな注目を集めている.非膜オルガネラには,カハール体以外にも核小体,核スペックル,パラスペックルなどの核内構造体,ストレス顆粒,プロセシング顆粒,生殖顆粒,神経顆粒など細胞質の構造体が知られている.いずれも蛍光顕微鏡を用いて各マーカー因子の免疫染色によってドット状に検出できる2).こうした非膜オルガネラには,周囲の細胞内空間との明確な“仕切り”がないのにもかかわらず,確固たる顆粒状の構造物として存在している.これらの多くの非膜オルガネラには核酸(RNA/DNA)が含まれており,さらに含有アミノ酸組成が著しく偏ったドメインをもつタンパク質が集約されていることがわかってきた2)〜4).近年,非膜オルガネラがこれらの構成因子の液-液相分離(LLPS,またはその関連現象)とよばれる物理現象によって形成されることが明らかになった.こうした相分離現象は,クロマチン構造形成,遺伝子発現制御,シグナル伝達,ストレス応答,オートファジーなどの細胞内制御機構,さらには神経変性疾患やコロナウイルス増殖経路に至るまで,さまざまな生体現象にかかわっていることが示されている.細胞内相分離は,混み合った細胞内空間を,膜のような仕切りを使わずに区画することによって,複雑な生命現象の正確な営みを支える基盤的機構といえる.
1.細胞内相分離はどのように引き起こされるか?
細胞内でみられる相分離現象は,しばしばサラダドレッシングのような状態に例えられる.つまり細胞内の液体空間は,タンパク質や核酸分子によって局所的に相分離形成された液滴が共存した状態と考えられる.そもそも相分離の研究は,物理学では古典的な研究分野であり,生体分子を含む柔らかい高分子ポリマー溶液中で観察される多彩な相分離現象を研究するソフトマター物理学という大きな研究分野に進展している.細胞内相分離も基本的にはこうした高分子ポリマーの相分離と同様の原理に基づいて誘発されると考えられる.タンパク質は,アミノ酸のポリマーでその配列や組成によって相分離を誘発する高分子ポリマーとして働いている.一般的に,タンパク質はアミノ酸配列によって決められた構造にフォールディングして機能するが,その一方で特定のアミノ酸の含有率が著しく偏って決まった構造をとりにくい天然変性領域をもつタンパク質が数多く知られている.特に非膜オルガネラを構成するタンパク質の多くは天然変性領域をもっていることが古くから知られていた2)〜4).そしてこの天然変性領域同士の多価相互作用によって相分離が引き起こされ,外部から隔てられた相分離空間となったものが非膜オルガネラとして働いていることが示された(図1).こうした天然変性領域のフラフラと構造をとらない状態は高分子ポリマーと類似しており,同様の原理に基づいた細胞内相分離が起こっていると考えられる.非膜オルガネラの構成タンパク質の多くは,天然変性領域とともに球状ドメインを併せもっている場合が多く,天然変性領域を介して形成した相分離空間内で,球状ドメインによる特定の機能を果たしていると考えられる.一方では,タンパク質の全長にわたって天然変性領域のまま働く超天然変性タンパク質ともよぶべき一群のタンパク質が最近発見され,多彩な生理現象にかかわっている可能性が浮上してきた5).またタンパク質以外にも,RNAが上記の多価相互作用に介入して相分離を促進したり,場合によっては抑制することも報告されている(図1).細胞内相分離は,温度などの周囲の環境や翻訳後修飾,また制御因子によって柔軟に制御されており,生体分子の振る舞いに秩序を与え,それを通して多彩な生命現象を駆動していることが明らかになりつつある.本書の第1章では,相分離を引き起こし制御するために働く因子の基本的な知見を相分離の原理とともに紹介する.
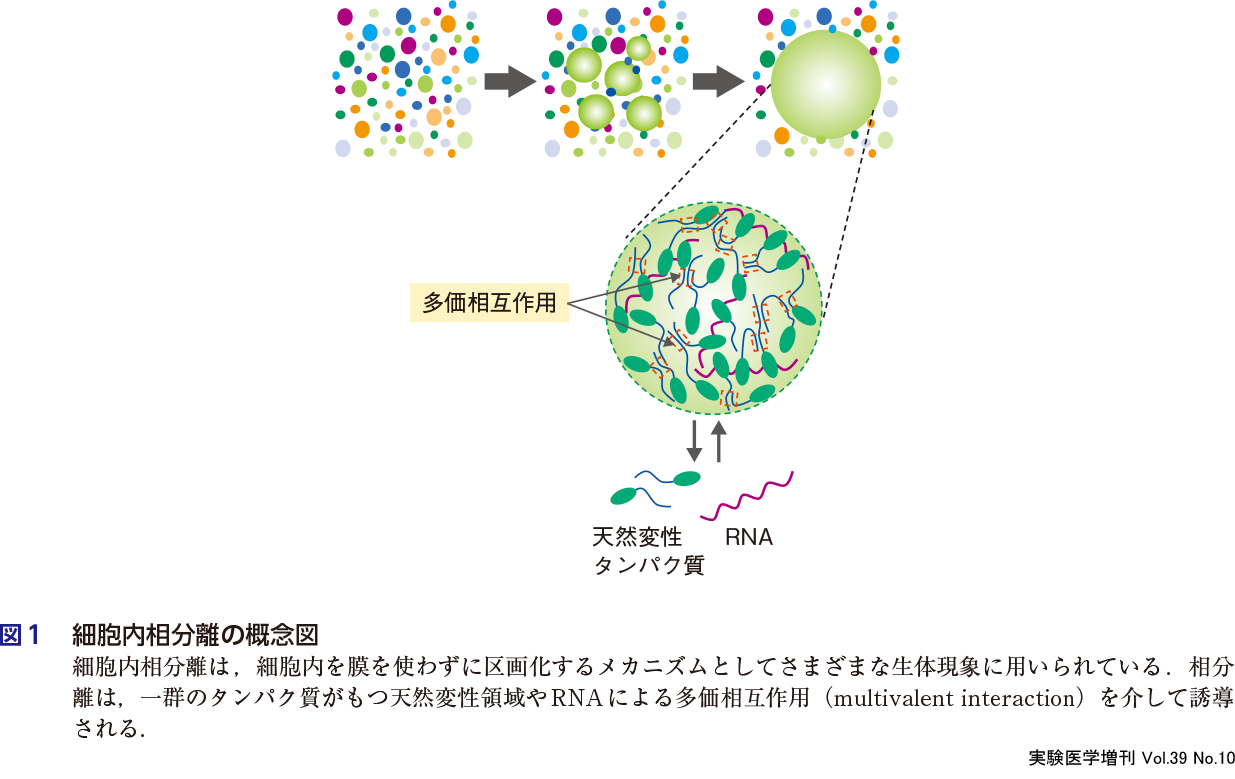
2.細胞内相分離によって何がつくられるのか?
細胞内相分離研究の先駆けとなった非膜オルガネラは,古くから光学顕微鏡観察によって観察されていた.その後,マーカータンパク質やRNAに対する蛍光プローブを用いた観察によって,次々と新しい構造体が発見されている(図2).細胞生物学者たちは,オルガネラの構成因子がダイナミックに内外を行き来していること,オルガネラが頻繁に融合/分離し液滴のように振る舞うことに気づき,それが相分離研究の先駆けとなった2)〜4).多くの非膜オルガネラにはRNA分子が含まれており,それと結合するRNA結合タンパク質がもつ天然変性領域が相分離を誘発する活性をもつことが,試験管内での液滴形成やハイドロゲル形成実験によって示された.またそうしたオルガネラのなかには,特定のRNA分子によって相分離形成が先導されるものもあり,そうしたRNAには天然変性タンパク質を選択的に集約させる配列が含まれている6).またRNAは相分離したオルガネラのサイズを規定したり,内部の微細構造をデザインする働きがあることも明らかになっている.RNAに依存した非膜オルガネラの多くは,RNA遺伝子のゲノム座位近傍に転写と共役して形成され,RNAの転写のオン・オフによってオルガネラの消長がコントロールされている.このような一過的な非膜オルガネラの形成は,周囲のクロマチンの配置に影響を与えることも知られていることから遺伝子発現制御への関与が想定されている.転写の活性化では,転写開始部位から比較的遠位に存在するエンハンサー領域に結合したコアクチベーターやメディエーターといった天然変性領域をもつ転写活性化因子が転写開始を促進している7).複数のエンハンサーが局所的に集まった領域は,特にスーパーエンハンサーとよばれ,そこにはさらに多くの転写調節因子やその領域由来のエンハンサーRNAが集まり相分離を引き起こし非膜オルガネラを形成して,複数のゲノム領域の転写活性化を担っている8).一方で,転写の不活性化に伴うヘテロクロマチン化でも,不活化領域の修飾ヒストンに結合する制御タンパク質の相分離がヘテロクロマチン化に重要な役割を果たしていることが報告されている9).このように,RNAやDNAに結合する制御タンパク質の多くが天然変性領域をもっていることは,相分離現象が遺伝子発現経路のさまざまな場面で重要な役割を演じていることを示唆している.本書の第2章では,これまで知られている数多くの非膜オルガネラの構造と機能について各論として紹介し,相分離を介して形成される細胞内構造に関する最新の知見を広く提供する.
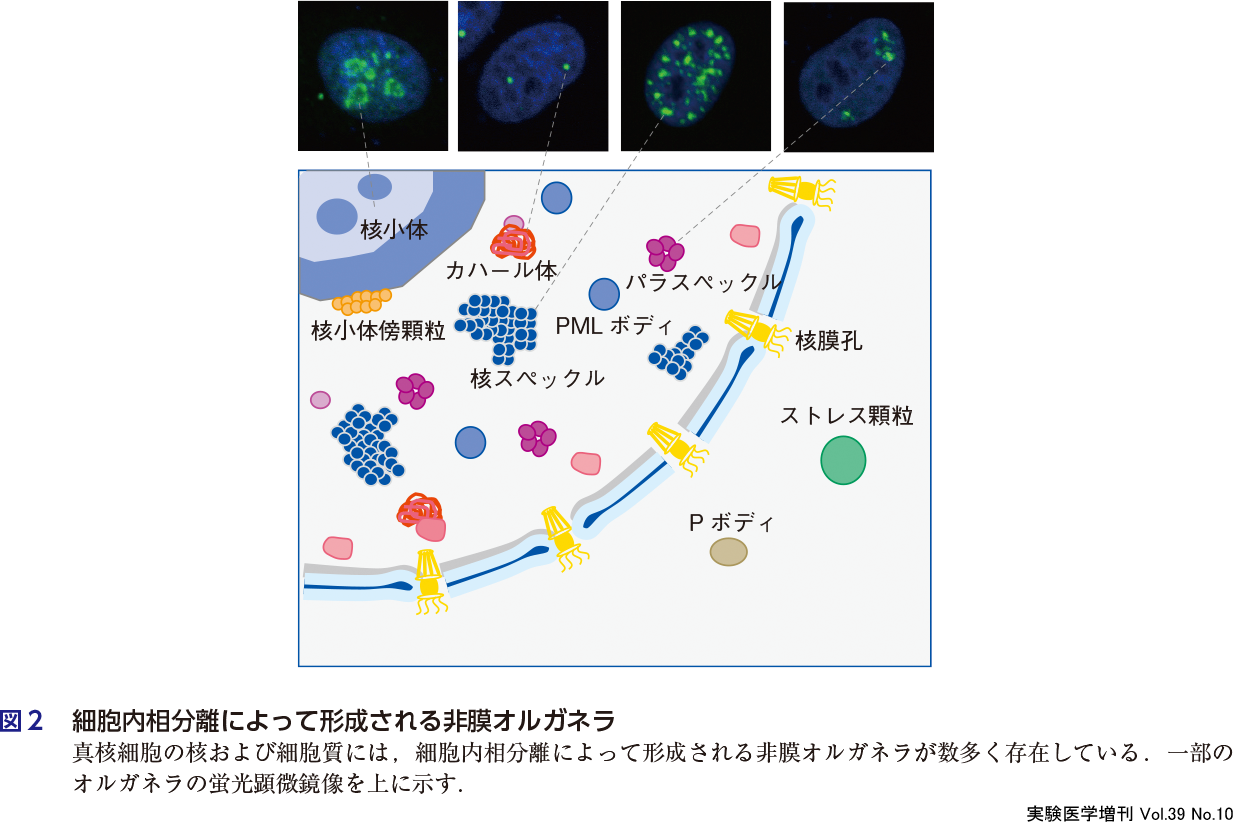
3.細胞内相分離にはどのような意義があるのか?
細胞内で相分離空間を形成することには,どのような意義があるのだろうか? 非膜オルガネラの機能としては,古典的な核小体でのリボソーム生合成のような巨大分子装置の工場としての機能があげられる.こうした工場では一連の生化学反応が効率よく順序立てて行われる必要があり,それが相分離によって隔離された空間に高濃度に集められた因子が,さらに内部で層構造を形成することによって行われている.こうした働きは,別名「るつぼ機能」ともよばれる(図3)4).一方で,特異的な因子を選択的に相分離空間に係留することによって,それらの因子が果たしていた機能を抑える働きも知られており,「スポンジ機能」とよばれている(図3)4).さらに核内では,非膜オルガネラはクロマチン領域の間隙に存在しているが,実際にクロマチンと相互作用することによって特定のクロマチン領域の配置や活性に影響を与える「ハブ」としての働きも提唱されている(図3)4)8)9).こうした分子機構に関する知見に加え,相分離現象がさまざまな細胞レベルおよび個体レベルでの生理現象に広くかかわっていることを示す知見が次々と報告されている.RNAやDNAがかかわる遺伝子発現だけでなく,タンパク質分解,細胞骨格重合,シグナル伝達,さらにはオートファジーといった多彩な細胞内現象にかかわる多くの因子が天然変性領域をもっており,それらを介した相分離現象がこれらの生体現象の特異性や効率などを制御していることが明らかにされている10).真核生物のタンパク質の30%以上は天然変性領域をもっているという報告もあり11),これらの多くが相分離現象を引き起こすとすれば,広範な生体内現象には,何らかの形で相分離がかかわっていることが予想される.本書の第3章では,最近明らかになってきた相分離を介して行われる生理現象についての知見を紹介する.
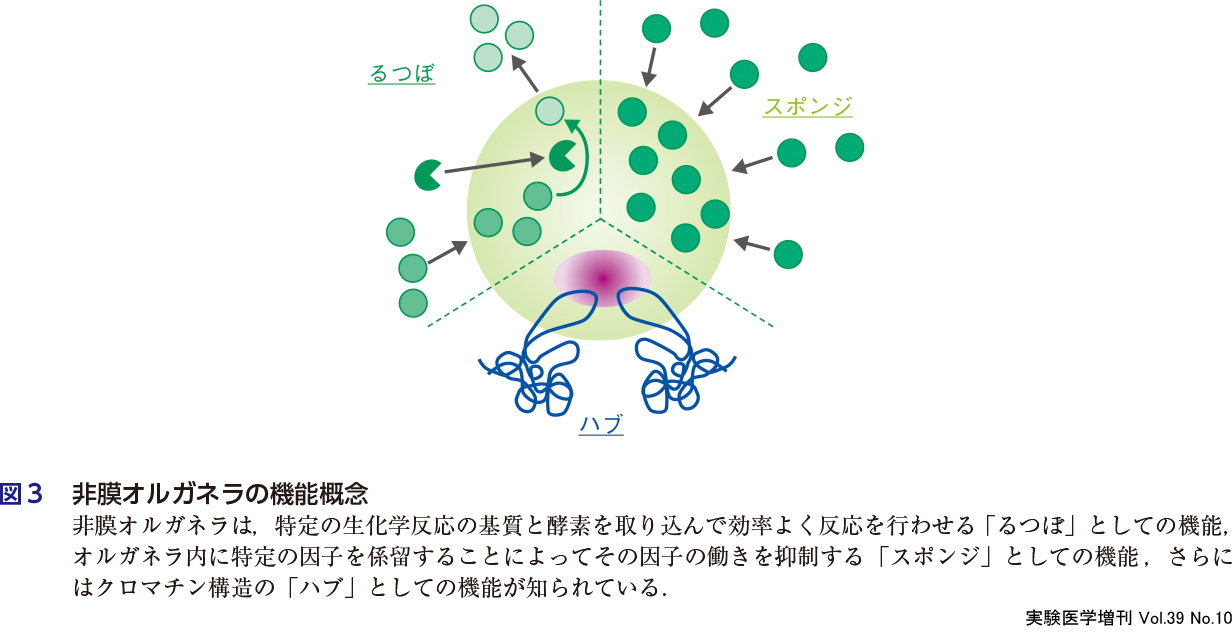
4.細胞内相分離の破綻は何をもたらすのか?
細胞内相分離の研究がこれほど注目を集めている理由の1つとして,疾患との接点が見出されたことがあげられる.特に難病として名高い家族性筋萎縮性側索硬化症(ALS)の原因タンパク質として同定されたTDP-43,FUS,HNRNPA1などのRNA結合タンパク質が共通して天然変性領域をもっており,相分離によって試験管内で液滴を形成すること,さらにその天然変性領域に疾患関連変異が同定されたことから,これらのタンパク質による相分離誘導と神経変性疾患の発症メカニズムとの関連が注目されるようになった10).これらの天然変性領域は,特にプリオン様ドメインともよばれ,試験管内での溶液組成,温度条件,時間経過によって液滴からアミロイド様凝集体に相転移することが示された(図4).さらに興味深いことに,ALSの病原性アミノ酸変異をもつFUSタンパク質の液滴は,野生型FUSの液滴よりも速く相転移することが示された.このことより,天然変性領域による相分離現象の異常によって毒性凝集体が形成され,それらが細胞内に蓄積するという分子経路が提唱された10).さらに最近では,神経変性疾患だけでなくがんや感染症の発症経路でも天然変性タンパク質による相分離現象の関与が報告されている.またコロナウイルス(SARS-CoV-2)の増殖経路に,ウイルスゲノムRNAとヌクレオキャプシッドタンパク質による相分離現象がかかわっていることが報告され注目を集めている12).相分離現象を標的とした創薬基盤研究も開始されており,相分離の視点での疾患研究はこれから発展する様相である.本書では第3章にこれらの最新トピックスを紹介する.
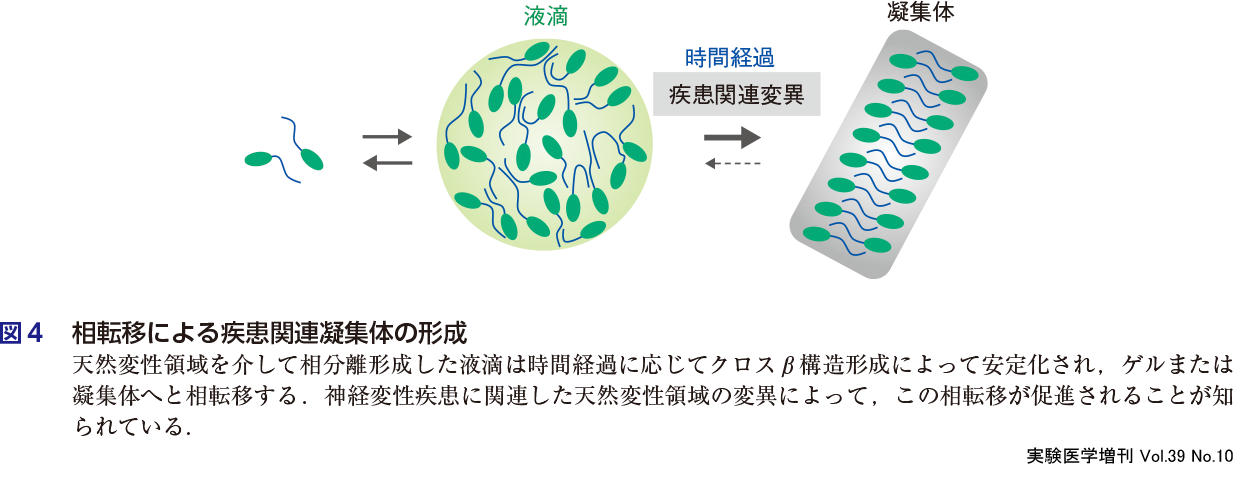
5.相分離をどのように研究したらよいか?
細胞内相分離の研究ははじまったばかりであり,多くの細胞生物学者が注目するようになった現在でさえ,細胞内における相分離現象を確実に捉えるスタンダードな解析法は確立されているとはいえない現状である13).細胞生物学者が手軽に採用する解析法としては,顕微鏡を用いてFRAPなどの既存の方法によって非膜オルガネラの構成因子の動態をモニターすること,相分離に関与する(一部の)分子間相互作用を阻害する試薬(例えば,1,6-ヘキサンジオール)の感受性を調べること,また試験管内においてタンパク質の液滴形成をモニターしそれにかかわるアミノ酸残基を解析することなどがあげられる.しかし,こうした解析だけでは,細胞内での相分離現象を正確に捉え,その機構を解析するには不十分である.特に,これからの細胞内相分離現象のさらなる理解には,細胞内での相分離体の挙動を定量的に計測する技術が不可欠である.そのために異分野の先端技術を生体分子の計測に適用する学際研究を積極的に推進することが,この研究分野の基盤をより確固なものにしていくうえで必須であろう.そこで本書では,第4章として相分離研究に威力を発揮すると考えられる先端計測技術を紹介する.
おわりに
混み合った細胞内空間で複雑な生化学反応経路が互いに混線することなく正確に進行するために,相分離による区画化が起こっているという考え方はとてもわかりやすい.また非膜オルガネラは,シードとなる分子の発現や修飾によってその形成を柔軟にコントロールでき,一過的に細胞内のピンポイントで相分離空間を形成し,特定の生化学反応を重点的に進行させることも可能である.また一連の生化学経路がオルガネラ内部の異なる区画やオルガネラ間を伝達しながら進行していくこと,またその分子の受け渡しに方向性を与える分子機構も存在することもわかってきた14).これまでに研究が進んでいるのは,光学顕微鏡で検出できるマクロな相分離構造体であるが,より微小な液滴が細胞空間には無数に存在し多岐にわたる反応経路を取りもっている可能性もある.この数年間で,細胞内相分離に関する論文が多数発表されたことにより,相分離研究は細胞生物学のホットトピックスへと急成長してきた.しかしその一方で,不十分な検証データだけで安易に相分離と結論づけているものが散見されることから,「猫も杓子も相分離」といった現状には警鐘が鳴らされている15)16).細胞内での定量的データに基づいた相分離現象の解析が欠落していることが指摘されており,すでによく研究されている非膜オルガネラのなかにも,その形態や挙動からいわゆるLLPSでは説明がつかない機構によって形成されている可能性も指摘されている.本研究分野の隆盛は喜ばしく今後の発展が大いに期待される一方で,より厳密な学問体系の整備が今後の課題である.
文献
- Cajal SRY:Trab Lab Invest Biol, 2:129-221, 1903
- Banani SF, et al:Nat Rev Mol Cell Biol, 18:285-298, 2017
- Kato M, et al:Cell, 149:753-767, 2012
- Shin Y & Brangwynne CP:Science, 357:doi:10.1126/science.aaf4382, 2017
- Tsuboyama K, et al:PLoS Biol, 18:e3000632, 2020
- Yamazaki T, et al:Cold Spring Harb Symp Quant Biol, 84:227-237, 2019
- Cho WK, et al:Science, 361:412-415, 2018
- Sabari BR, et al:Science, 361:doi:10.1126/science.aar3958, 2018
- Sanulli S, et al:Cold Spring Harb Symp Quant Biol, 84:217-225, 2019
- Alberti S & Hyman AA:Nat Rev Mol Cell Biol, 22:196-213, 2021
- Ward JJ, et al:J Mol Biol, 337:635-645, 2004
- Iserman C, et al:Mol Cell, 80:1078-1091.e6, 2020
- Alberti S, et al:Cell, 176:419-434, 2019
- Hondele M, et al:Nature, 573:144-148, 2019
- McSwiggen DT, et al:Genes Dev, 33:1619-1634, 2019
- Leslie M:Science, 371:336-338, 2021
著者プロフィール
廣瀬哲郎:1995年名古屋大学大学院理学研究科にて博士取得,その後,名古屋大学助手,米国イェール大学ポスドク,東京医科歯科大学特任准教授,JSTさきがけ専任研究員,産業技術総合研究所グループ長,北海道大学教授を経て,2020年より大阪大学大学院生命機能研究科教授に就任した.この間植物から動物までのさまざまな生物種でRNAが織りなす多彩な現象に触れてきた.現在は哺乳類ゲノムから合成されるノンコーディングRNAが相分離を介して細胞内現象を制御する分子機構について,手垢のついていない涼しげな研究をめざしている.