
概 論
シン・骨格筋ワールド:
<
Shin・skeletal muscle world:<— skeletal muscle research rises to a crescendo
藤井宣晴
Nobuharu L Fujii:Department of Health Promotion Sciences, Graduate School of Human Health Sciences, Tokyo Metropolitan University(東京都立大学 人間健康科学研究科 ヘルスプロモーションサイエンス学域)
はじめに─骨格筋研究の潮流:序
骨格筋は動きを生む生体内器官であり,筋収縮がその源になっている.筋収縮によって,より高次の動作や運動を実現するためには,中枢神経からのコマンド発信と,それを伝える運動神経の働きが必須となる.生命体は,中胚葉由来の骨格筋と,外胚葉由来の神経系という,発生上,属性が異なる細胞同士のマッチングを膨大な時間をかけて試み,両者の協同作業を可能にした.すなわち「動き」の獲得は,生物の進化の方向を決定する要因の1つと考えられる1)2).いまだ不明の,おそらくは複数の複雑な過程を経て,ヒトは直立姿勢をとり,二足歩行が可能となった.姿勢維持や歩行から解き放たれた両腕は,指を器用に操る繊細な技を獲得し,より高度な生活様式を得た結果として,大脳新皮質を発達させた3).
筋収縮とは,細胞内Ca2+の上昇をトリガーに4),ミオシン線維の束の中にアクチン線維が滑り込む現象である5).骨格筋の収縮機能が健全に保たれていると,わたしたちは自由に暮らすことができるが,筋難病や事故でその機能が損なわれると,生活の質(quality of life,QOL)が低下するだけではなく,場合によっては命の危険にもさらされる.そのため,これらの治療・予防法の確立は急務であり,研究者の莫大なエネルギーが注がれている6).また,加齢による筋量および筋力の低下(サルコペニア)が,身体の脆弱・機動性の欠如・障害・死亡のリスクを増大させることが明らかとなり,超高齢社会を迎えた日本では,サルコペニアへの対策に重点がおかれている7).
1.骨格筋の機能と量を保つ意義:破
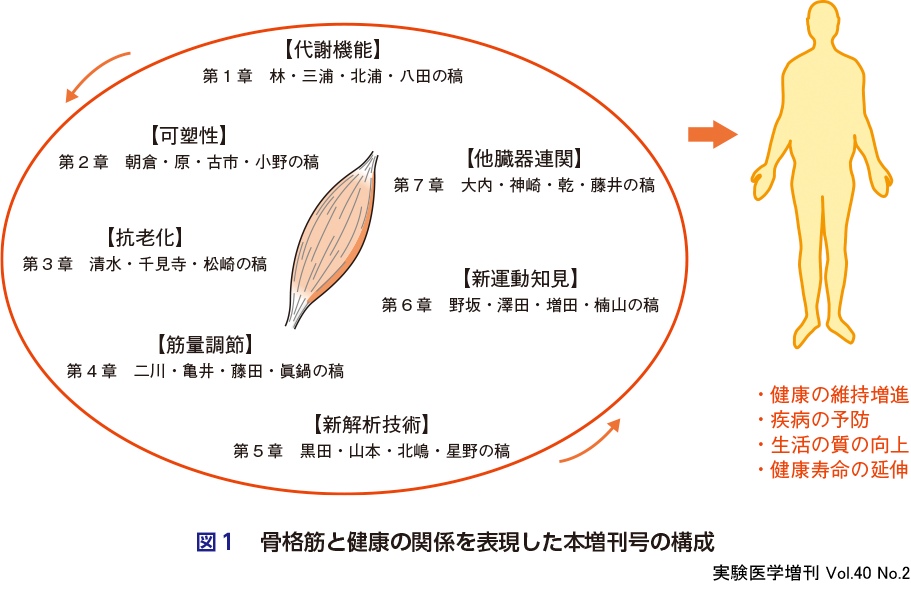
前項で述べたような骨格筋研究の潮流は,骨格筋の機能と量を維持すべきことを示している(図1).これらが阻害あるいは減少すると,行動が制約され,QOLが低下し,活動量とエネルギー消費量が減少して,疾病の発症や支援・介護の導入へと事態が進行してしまう.一方,健康長寿が,行動の制約とは別の原因ではばまれるケースも示唆されている.むしろ,機能性を維持した骨格筋を適正量もつこと自体が,健康増進,疾病予防,あるいは生命力維持に重要であるとの示唆である.
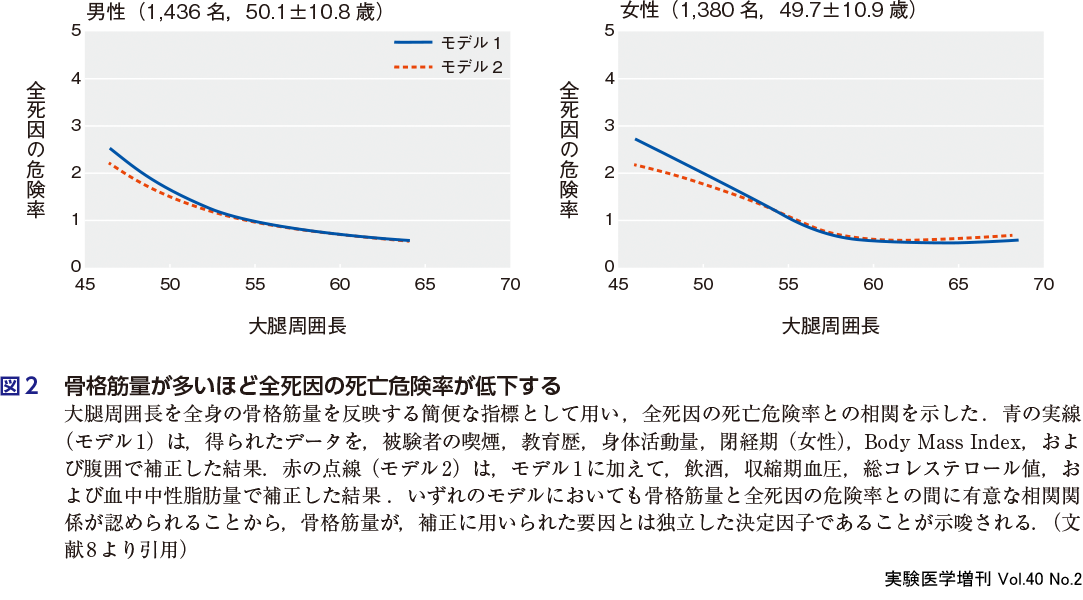
Heitmann BLらは,がん,冠動脈疾患,および脳梗塞のない30〜60歳の者を対象に,全身の筋量を簡便にあらわす指標として大腿の周囲長を測定した.その後,約15年間の追跡調査を行い,すべての死因における死亡の危険率との相関を検討したところ,大腿が太い者ほど危険率が有意に低いことが判明した(図2)8).この知見は,筋量の多い者ほど死亡リスクが低いのは広い年齢層に普遍的で,高齢者のサルコペニアに限られないことを示唆している.
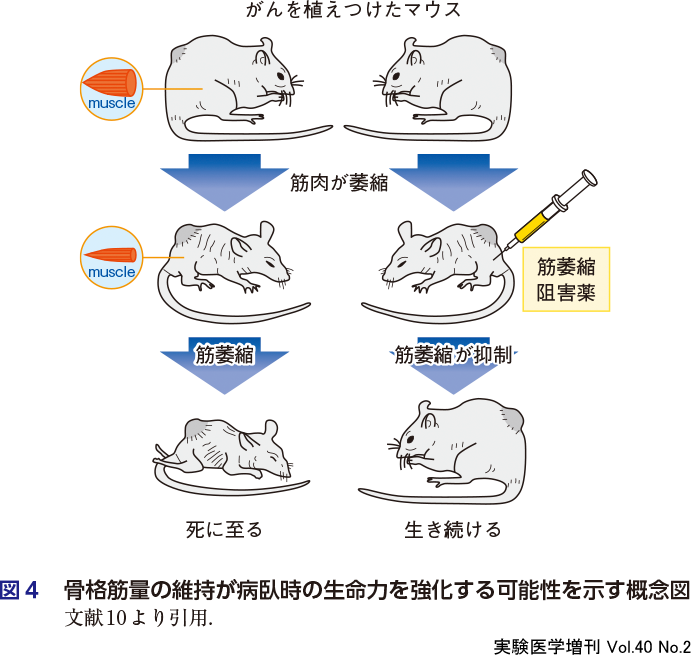
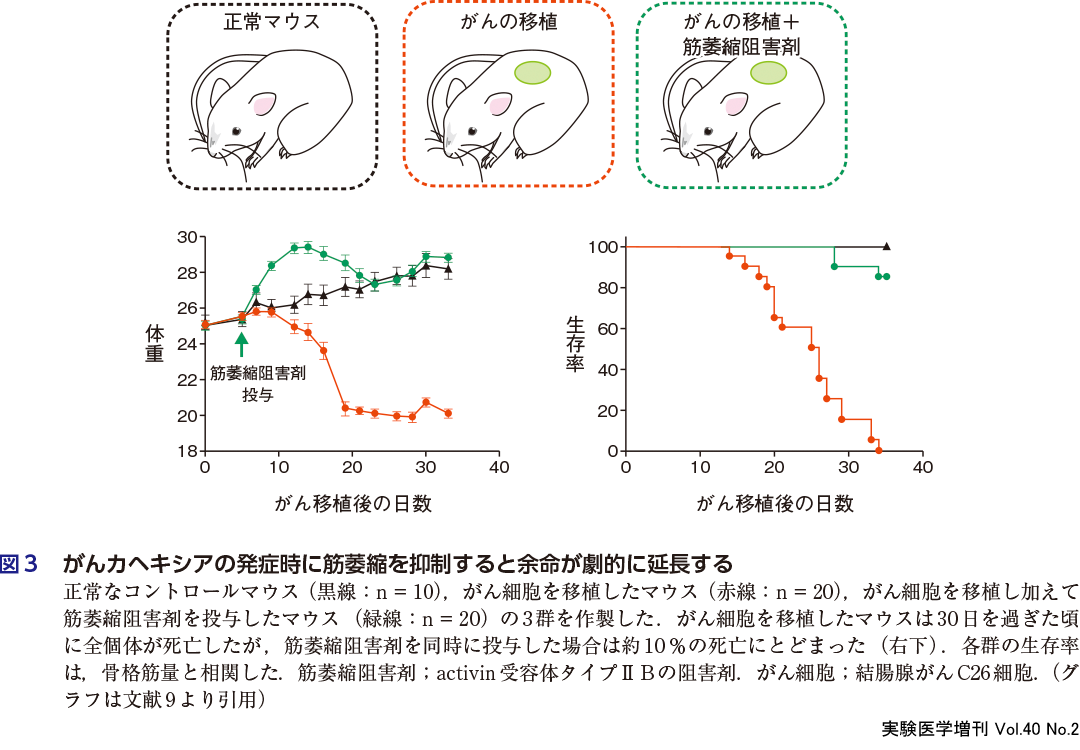
カヘキシア(悪液質)とは,がん,腎不全,あるいは糖尿病などの慢性疾患が原因で生じる複合的代謝異常のことである.2000年代に入り,カヘキシアがQOLや生命予後に悪影響を有することが明らかになり,注目されるようになった.カヘキシアは骨格筋の急激かつ大量な減少を特徴としているが,それとQOLや生命予後に対する悪影響との関係については不明であった.Zhou Xらは,骨格筋の成長を抑制する液性因子であるマイオスタチンの作用に注目し,実験を行った9).結腸腺がんC26細胞を移植されたマウスは,移植から30日が過ぎた頃に全個体が死亡した(図3右下).死亡時の体重は,移植のない正常マウスと比較して35%減少しており(図3左下),その主因は除脂肪体重,すなわち骨格筋の減少にあった.つぎに,がん細胞を移植したマウスに,マイオスタチンの受容体であるactivin受容体タイプⅡBの阻害剤を,筋萎縮阻害剤として投与した.すると,体重の減少は生じず(むしろ一過性に増加し)(図3左下),生存率が劇的に改善された(図3右下).これらの知見は,骨格筋の存在自体が生命力の維持にかかわっていることを示唆する(図4)10).しかしそのメカニズムは不明のままで,今後解き明かさねばならない課題である.
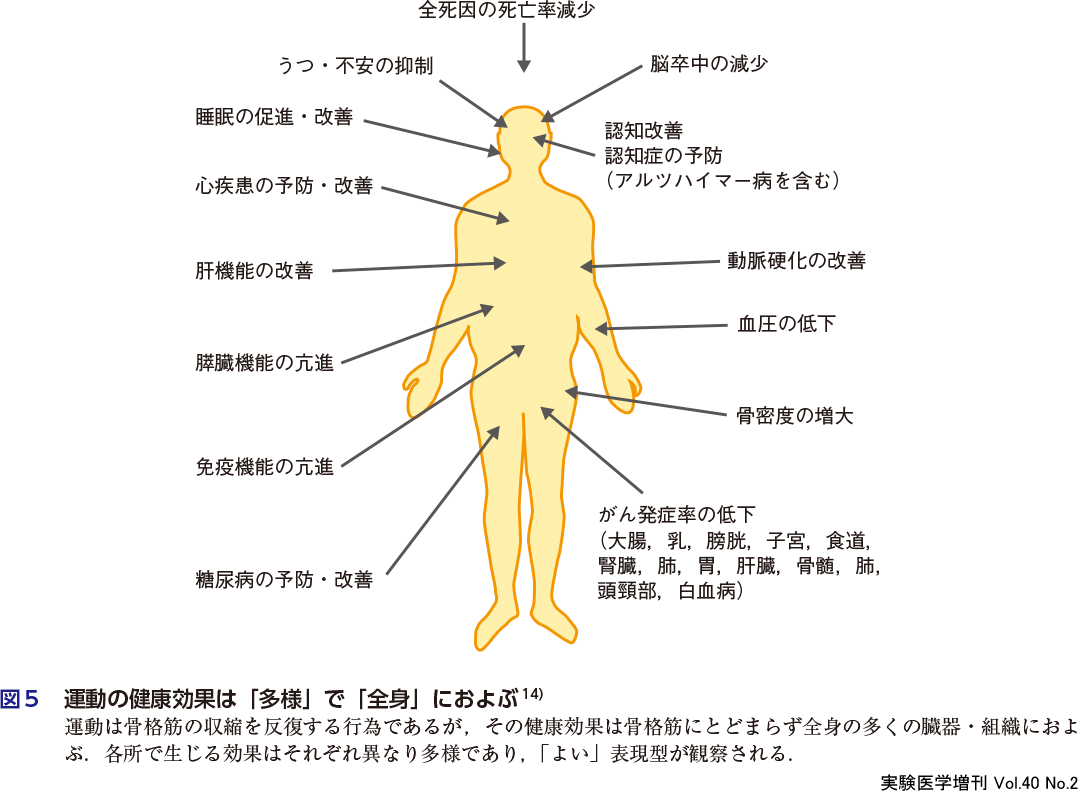
骨格筋の機能と量を適切に保つ方策に,運動がある.運動は,骨格筋のインスリン抵抗性を予防・改善する.インスリン抵抗性とは,インスリンへの感受性が低下し,その作用が十分に生じない状態のことで,2型糖尿病の誘因とされる.糖尿病は,高血圧・高脂血症・肥満とともに,動脈硬化を進行させる危険因子である.一方で,インスリン抵抗性は糖尿病だけではなく,高血圧・高脂血症・肥満の,すべての危険因子の誘因とする考えも提唱されている11).これに従えば,インスリン抵抗性の抑制で多くの疾患を予防できることになる.骨格筋をダイナミックに収縮させる運動は,それが有酸素運動でもレジスタンス運動〔標的とする筋肉に抵抗(レジスタンス)をかける,いわゆる筋力トレーニング〕でも,健常者ではインスリン感受性を高め,すでにインスリン抵抗性がある者ではそれを改善させる12).単回の運動セッションでは,運動中に血液中のグルコースが骨格筋に取り込まれるとともに,運動終了後には一過性にインスリン感受性が高まる.さらに,運動をある期間継続すると,インスリン感受性はさらに亢進し,かつ定着する.このように,運動によって骨格筋のインスリンに対する感受性を高め抵抗性を抑止することで,多くの疾患への罹患リスクを低減できる可能性がある.図5に示すように,運動は「多様」な運動効果を「全身」にもたらしてくれる13).骨格筋と健康の因果関係は,いまだ多くのことが不明だが,最新の研究はいくつかのベールを確実にはがしてきた.
2.骨格筋と健康の関係:急(図1)
1)骨格筋の代謝調節を支える機序(第1章)
生命を維持するのに必要なエネルギーは,各細胞内のアデノシン三リン酸(adenosine triphosphate,ATP)内に保持されている.骨格筋は安静時のATP消費量が最も多い器官で,運動時にはその量が飛躍的に増す.しかしATP分子の数は基礎代謝を1分間支える程度しかなく,消費しながら同時に合成も行う代謝調節がなされる.超高齢社会を迎えた日本においては,代謝機能を80年以上も健全に維持しなければならず,これからの研究の重要性が増している.
運動中に骨格筋はインスリン非依存的に細胞内へグルコースを取り込み,終了後は長時間にわたってインスリン感受性が高まる(第1章-1).脂質分子は保持できるエネルギー量が多く,ATP合成に効率的であるが,運動トレーニングを継続的に行うと,脂質の利用能力が向上する.さらに,脂質を構成する脂肪酸や極性頭部の種類の違いが,骨格筋機能に影響するため,脂質クオリティ研究の展開が注目されている(第1章-2).タンパク質・アミノ酸は,骨格筋を構成する主要因子であるだけでなく,栄養不足状態ではエネルギー供給源ともなる.分岐鎖アミノ酸は,骨格筋がエネルギー源として利用できる唯一のアミノ酸だが,その代謝は骨格筋内で完結しているのではなく,全身の代謝や,糖・脂質代謝とカップルしているという(第1章-4).乳酸は疲労物質として,長く悪者あつかいされてきた.しかし最近では,解糖系と酸化的リン酸化系をつなぐエネルギー基質として認識されるようになっている.くわえて,シグナル伝達物質として働くことも示唆されており,これまでとは異なる側面が見えはじめている(第1章-4).
2)骨格筋機能を変容させる外部/内部環境因子(第2章)
可塑性とは,おかれた環境に応じて,形,容積,機能を変える能力・性質のことである.傷害を受けた後に再生・回復する能力もこれに含まれる.骨格筋は可塑性に富んでいて,損傷から再生回復する能力が高く,筋の使用頻度に応じて肥大や萎縮する.骨格筋の高い可塑性が対応する外部/内部環境因子は複数あるが,環境因子の変容が過度に激しい場合は,適応機構が破綻してしまう場合もある.
サテライト細胞は骨格筋の体性幹細胞で,ふだんは細胞分裂を停止した休止状態にあるが,何らかの刺激が入ると増殖・分化し,筋量の増大や筋損傷の回復に貢献する.刺激には機械的刺激,液性因子,筋損傷など複数あるが,それらの信号を適切に受容するためのニッチが重要であることがわかってきた(第2章-1).さらに,サテライト細胞が骨格筋細胞に融合したり,筋が損傷を受けたりした際の,細胞膜の修復・再生機構が,骨格筋の恒常性には欠かせないことが指摘され,細胞膜のクオリティ調節の理解の必要性が強調されている(第2章-2).グルコースは多くの細胞にとって重要なエネルギー供給源であるが,骨格筋細胞の増殖能力に関しては逆で,低グルコース環境の方が増殖は亢進することが明らかになった(第2章-3).一方,骨格筋細胞の特性や機能には身体部位特異性があって,これは筋線維タイプの違いでは説明しきれないという.そこで,内部環境因子として,身体位置記憶という新概念が案出されている(第2章-4).
3)個体の老化を支配する骨格筋の老化(第3章)
個体の老化とは,生まれてから死に至るまでの時間経過(加齢)に伴う,身体機能の低下をさす.これに対して細胞老化とは,細胞の増殖が不可逆的に停止した状態をさし,テロメア短縮によるヘイフリック限界,DNA損傷,がん遺伝子の活性化などによって誘導される.細胞老化は個体老化の一要因と考えられるが,一方で損傷したDNAの複製を防ぐことでがん化を抑制しているとも考えられる.
筋収縮時には,ミトコンドリアで産生された活性酸素種がDNA損傷のリスクを高めるため,酸化ストレスは骨格筋細胞を老化させる一因と考えられている.しかし,抗酸化処置が骨格筋の恒常性維持に必ずしも好影響をおよぼすとは限らず,複雑な調節機構の存在が予想されている(第3章-1).運動は老化に抗する有効策とされるが,実際には細胞老化の刺激因子も活性化させる.この事実は,運動が骨格筋にもたらす現象を考える際に,有益な老化と有害な老化を見極める必要性を示す(第3章-2).また,骨格筋細胞だけでなく,サテライト細胞も老化することが知られており,サルコペニアへの関与について知見が集積しつつある(第3章-3).
4)骨格筋の肥大と萎縮(第4章)
骨格筋は可塑性に富むため,機械的負荷をかけると肥大するし,かけないと萎縮する.そのダイナミックな変化を可能とする機序の理解が,骨格筋の機能と量を調節するための方策を確立するためには欠かせない.
骨格筋に負荷がかからない状態で生じる萎縮は廃用性筋萎縮とよばれ,その主因に酸化ストレスがあげられている(第4章-1).細胞老化との連関が,今後の新たな研究分野になりえるだろう.筋量の維持・増進にはレジスタンス運動が推奨されるが,食事によるタンパク質摂取も同様に重要である.タンパク質を摂るだけで,同化作用は高まるが,その効果が残る時間は短いので,運動と組合わせることで効果的な筋量増大が可能という(第4章-2).さらに,絶食すると骨格筋細胞では飢餓応答が起こり筋萎縮を進行させるが,その細胞内情報伝達ネットワークが明らかにされてきている(第4章-3).一方,培養細胞を用いた研究では筋張力を測定することができないため,筋細胞の横幅の長さを計測して,筋肥大の目安とすることが多い.ところが実際は変化量が小さく,実験間の誤差やバイアス混入なども生じやすいのが問題であった.最近になって,工学と細胞生物学の手法との融合で,筋張力の近似的定量が可能となった(第4章-4).
5)骨格筋の秘めた特徴を捉える新技術(第5章)
研究では,新しい技術の導入を怠ると,大きな展開が難しくなる.研究の進展は新鮮なアイディアとともに新技術の登場にも支えられている.
骨格筋「細胞」の収縮は,運動神経からの入力があれば全力で収縮し,入力がなければ収縮しないという,「0か1」しかとらない全か無の法則に従う.しかし,実際にはわたし達は筋力をさまざまに調節できる.その理由は,筋組織を構成する個々の細胞が,閾値の異なる運動神経によって支配されており,中枢神経からの指令強度が低いと収縮する細胞数が少なく,高いと多くなるためである(運動単位の動員増大).骨格筋「組織」の収縮強度の調節は,古くから運動単位の動員で説明されてきた.ところが最近,より細やかな張力調節を行う機構の存在が示された.従来の方法では,収縮刺激に対する骨格筋細胞応答のばらつき(変動)を,細胞内変動と細胞間変動に分離して計測できなかったが,新技術を用いてそれを可能にし,応答多様性効果を考慮した情報数理のモデル解析を行ったところ,これまでノイズとして扱われてきた応答のばらつきは制御情報として働いており,細胞間変動によって刺激強度の違いを細かく区別できるようになるために,組織レベルでは,かえって正確な組織応答制御が可能になるという(第5章-1).筋収縮に必要なATPの重要性を示す過去の研究結果の多くは,生化学や薬理学的に導かれたものである.現代では,生きたマウスの生体内で,細胞内ATP動態を可視化することで証明するという,新たな角度からの試みが進んでいる(第5章-2).このような遺伝子組換えマウスの創出技術は,医学・生命科学の進歩に大きな貢献をしてきたが,その進化はいまも続いている(第5章-3).オミクス解析も進化を続ける分野の1つである.骨格筋では,遺伝子発現・シグナル経路・代謝階層にまたがるトランスオミクスネットワークの構築・解析により,これまで見落とされていた,ペントースリン酸経路の新たな作用が明らかになった(第5章-4).
6)骨格筋と運動の新たな関係の顕在化(第6章)
骨格筋と運動との関係は,古くから研究されてきた.現在も両者の秘された関係は堀り尽くされておらず,興味深い発見が続いている.
運動は筋収縮の反復・持続で成り立っているが,筋収縮の様式には,アイソメトリック,コンセントリック,およびエキセントリックの3種類がある.最近は,筋が伸張しながらも力を発揮するエキセントリック収縮が注目されており,筋力増大だけではなく,さまざまな疾病予防にも効果が認められている(第6章-1).筋収縮がもたらす健康の恩恵効果は複数の機序により生じている.収縮によって生じる力学的刺激も主要な役割を担うが,その端緒が筋組織の内圧変化によって生じる間質液の流動にあることが明らかにされた(第6章-2).このことは,細胞レベルの研究だけでは運動の本質に迫ることができないことを指摘している.ミトコンドリアでのATP合成には酸素分子が必要で,その供給を骨格筋で媒介しているのがミオグロビンである.漠然と,ミオグロビンの局在は細胞質内と考えられてきたが,じつはミトコンドリア内に局在し,呼吸鎖複合体の一部と直接的な相互作用を有する可能性が示された(第6章-3).妊娠期に妊婦の生活習慣が乱れると,生まれた子どもが成人した際に慢性疾患発症のリスクが高まるという仮説が,近年になって散見されていた.その仮説を支持する報告がなされた.妊娠マウスに運動をさせると,母体からある液性因子が分泌され胎盤を刺激し,さらに胎盤から胎児側に液性因子が分泌されて,母体の運動効果が胎児に伝達されるという(第6章-4).
7)骨格筋と他臓器の連関(第7章)
現在は,各臓器の役割を臓器単独で理解するのではなく,生体内に存在する臓器間ネットワーク上で担う役割を理解するのが本流となっている.骨格筋についても,他臓器との連関および異種機能との連関が,研究の重要課題になっている.
骨格筋から分泌される生理活性物質は,総称してマイオカインとよばれ,他の臓器とのコミュニケーションに働くとする仮説がある.しかしマイオカイン研究の歴史はまだ浅く,骨格筋から分泌されていることを証明する条件(骨格筋特異的に発現量を増減させるなど)が整っていない報告があることも指摘されている.これらの条件を整えた確度の高い研究成果が日本からは発信されている(第7章-1).また,骨格筋細胞が免疫細胞と連関するニッチが,筋組織内の毛細血管にあり,そこでは生活習慣病の増悪因子として働くサイトカインが,良性作用を発揮していることが明らかにされた(第7章-2).骨格筋は腱と骨との連関で動作を生じさせているが,3者の連関は,それら近傍にある複数の異種細胞間との,さらなるコミュニケーションで成り立っているという(第7章-3).これら骨格筋の他臓器連関の研究は,発展が期待される分野であるがゆえ,注意深く進める必要がある(第7章-4).
おわりに─シン・骨格筋ワールド:<
骨格筋の研究は,萌芽からしだいに多様性が加わり,それにかかわる研究者も増え,成果の蓄積にも厚みができてきていると感じる.各章の見出しに付した「序」「破」「急」は雅楽の楽曲を構成する3つの楽章のことで,序ははじまり,破は展開,急は高まりをあらわす.高まりをみせている現在の骨格筋研究は,今後も勢いをおとすことなく進展していくだろう.そのことを本概論のタイトルで,「徐々に強く」を意味する音楽記号のクレッシェンドになぞらえ表現した.そのようななか,骨格筋研究の先端で活躍される26名の気鋭の先生方に協力いただくことができたのは,またとない喜びである.おかげで,独自のアイディアと新技術の創出のトピックで彩られた,魅力的な構成になったと思う.本増刊号をきっかけにして,多くの方に骨格筋への興味をもっていただけたら,幸いである.
行こう! シン・骨格筋ワールドへ.
文献
- Jékely G:Proc Biol Sci, 278:914-922, doi:10.1098/rspb.2010.2027(2011)
- Bramble DM & Lieberman DE:Nature, 432:345-352, doi:10.1038/nature03052(2004)
- 「Restless Creatures-The Story of Life in Ten Movements」(Wilkinson M), Hachette Book Group, 2016
- Ebashi S & Ebashi F:Nature, 183:687-688, doi:10.1038/183687b0(1959)
- Huxley H & Hanson J:Nature, 173:973-976, doi:10.1038/173973a0(1954)
- 「超高齢社会に挑む骨格筋のメディカルサイエンス」(武田伸一/編), 実験医学増刊 Vol.36 No.7, 羊土社, 2018
- 「特集:骨格筋の維持機構を暴き、サルコペニアに挑む!」(上住聡芳/企画), 実験医学 Vol.38 No.16(2020年10月号)羊土社, 2020
- Heitmann BL & Frederiksen P:BMJ, 339:b3292, doi:10.1136/bmj.b3292(2009)
- Zhou X, et al:Cell, 142:531-543, doi:10.1016/j.cell.2010.07.011(2010)
- Leading Edge. In This Issue. Combating cancer cachexia:Cell, 142:531, 2010
- Alberti KG, et al:Lancet, 366:1059-1062, doi:10.1016/S0140-6736(05)67402-8(2005)
- Fujii N, et al:Am J Physiol Endocrinol Metab, 291:E867-E877, doi:10.1152/ajpendo.00207.2006(2006)
- Piercy KL, et al:JAMA, 320:2020-2028, doi:10.1001/jama.2018.14854(2018)
著者プロフィール
藤井宣晴:1994年,筑波大学体育科学研究科修了後,筑波大学応用生物化学系でポスドク.’99年,米国ジョスリン糖尿病センター.2008年,東京都立大学人間健康科学研究科.骨格筋の研究をはじめて20年あまり.最近は,本増刊号で紹介する分野だけではなく,機械工学,人工知能,緩和医療,理学療法,畜産・水産,サプリメント開発,美容,芸術など,思いもよらなかった分野と交流する機会が増え,楽しい.WANTED! Postdocs and Graduate Students Who Work Hard with Us.http://www.comp.tmu.ac.jp/muscle/