
概 論
栄養・代謝物シグナルと食品機能
―栄養学の歴史と食品機能の作用機序
Nutrition, metabolite signal and food function
亀井康富
Yasutomi Kamei:Laboratory of Molecular Nutrition, Graduate School of Life and Environmental Sciences, Kyoto Prefectural
University(京都府立大学大学院 生命環境科学研究科 分子栄養学研究室)
[略語]
- eNOS:
- endothelial nitric oxide synthase(内皮型一酸化窒素合成酵素)
- FAD:
- flavin adenine dinucleotide(フラビンアデニンジヌクレオチド)
- FXR:
- farnesoid X receptor(ファルネソイドX 受容体)
- GPCR:
- G protein-coupled receptor(G タンパク質共役型受容体)
- HDAC:
- histone deacetylase(ヒストン脱アセチル化酵素)
- KEGG:
- Kyoto Encyclopedia of Genes and Genomes
- LSD1:
- lysine specific demethylase 1(リジン特異的脱メチル化酵素1)
- LXR:
- liver X receptor(肝臓X 受容体)
- NAD:
- nicotinamide adenine dinucleotide(ニコチンアミドアデニンジヌクレオチド)
- PPAR:
- peroxisome proliferator-activated receptor(ペルオキシソーム増殖剤応答性受容体)
- RAR:
- retinoic acid receptor(レチノイン酸受容体)
- SAM:
- S-adenosylmethionine(S- アデノシルメチオニン)
- TCA:
- tricarboxylic acid(トリカルボン酸)
- TET:
- ten-eleven translocation(メチル化シトシン水酸化酵素)
はじめに
「私たちは,なぜ食べるのか?」
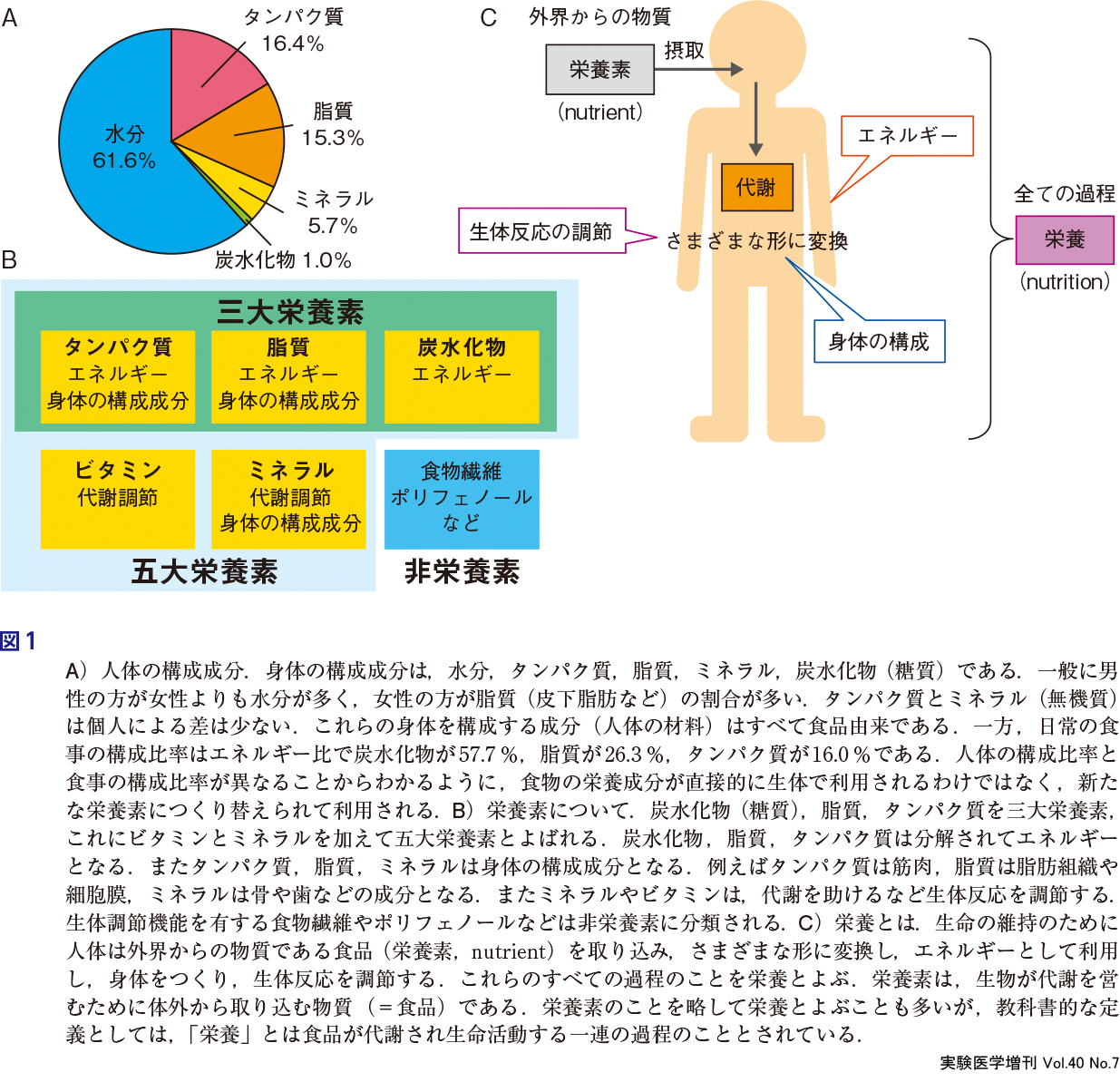
自動車にガソリンを入れて燃料にするように,ヒトは食品を摂取して燃料にする.それは半分正解だが,半分は不正解である.食品は身体をつくる材料になっている.これが自動車とヒトとの大きな違いである.食品は体内に取り込まれ,身体を構成する生体物質となり,代謝回転しながら常に入れ替わっている1).人体を構成する成分は,水分,タンパク質,脂質,ミネラル,炭水化物などである(図1A).これらの人体の材料はすべて食品由来である.また,後述するように,食品にはエネルギー以外の機能的な役割もある.
摂取された食品は消化・吸収され代謝を受ける.炭水化物,脂質,タンパク質は分解されてエネルギー源として利用される.またタンパク質,脂質,ミネラルは臓器,細胞,オルガネラなどの構成材料として身体の組織を形成する.さらに,ミネラルやビタミンは代謝を助ける働きをもっている.生命の維持のために,生体は外界からの物質(栄養素:nutrient)を摂取し,エネルギー,身体をつくる材料,生体反応の調節因子などさまざまな形に変換する(図1B).「栄養素」とは生物が代謝を営むために体外から取り込む物質のことであり,「栄養」とは食品が代謝され生命活動を行う一連の過程のことを指している(図1C)2).
本稿では,まず栄養学の歴史について簡単に述べ,そして食品機能の作用機序(メカニズム)に関して重要な,①核内受容体,②代謝物によるシグナル修飾,③食品成分と腸内細菌,および④代謝物研究の発展に必須の手法であるメタボロームについて整理し,概説する.
1.栄養学の発展
1)栄養学のはじまり
エネルギー代謝に関する発見が18世紀にラボアジェによって示された.生体は呼吸により酸素を使って燃焼を起こし二酸化炭素を発生させる.この呼吸の量は発生する熱量(エネルギー)に比例し,食物の摂取により増大することからエネルギー代謝の概念が築かれた.
20世紀になって栄養素の代謝研究がはじまり,1937年(昭和12年)にはクレブスにより,糖質が解糖され二酸化炭素と水へ酸化されてエネルギーを産生するTCA回路が発見された.さらに脂質も酸化されてエネルギー源となることが解明された.また脂肪(脂質)は単なるエネルギー源だけではなく,成長や生殖などの生理作用に欠かせない必須脂肪酸を含むことも明らかにされてきた.一方,タンパク質はアミノ酸から構成されていることが示された.その後,体内で合成されない必須アミノ酸と合成される非必須アミノ酸の発見,さらに各種タンパク質やアミノ酸の生理作用の研究へと発展していった(第1章-1).
2)栄養素の発見(欠乏症と過剰症)
コッホやパスツールらにより感染症の原因となる病原菌が数多く発見され,病気には対応する病原菌が存在すると信じられてきた.しかし,病原菌の見つからない難病が存在していた.白米を好んで食べる江戸の人々が発症した「江戸わずらい」という病気は,後に脚気であることがわかり,その治療には,米ぬかの成分が有効である.鈴木梅太郎によってオリザニンと名付けられたその成分はビタミンB1である.また,ヨーロッパの大航海時代には,新鮮な野菜や果物の摂取不足による壊血病が問題となっていたが,その原因がビタミンCの欠乏であることが発見された.このような脚気,壊血病に加えて夜盲症(ビタミンA)やくる病(ビタミンD)のような栄養素の欠乏による疾患の原因としてビタミンが発見され,これらのビタミンの発見に対して1930年代〜1940年代に複数のノーベル賞が授与された.さらにミネラルの欠乏症も発見され,その生理作用が明らかになってきた.近年,ミネラルの細胞機能のメカニズムの解析が進んでいる(第1章-3,4).また,ポリフェノールなど,これまで栄養素とは認識されていなかった食品由来成分(非栄養素とよばれる)が固有の生理活性をもち,健康や疾病予防に有効であることもわかってきた2).例えば,緑茶カテキンを感知するセンサーの存在が報告されている(第1章-7).このように食品由来成分のなかに人体の恒常性維持に必須の成分が存在することが明らかにされていった.すなわち,病原菌ではなく,食品中の微量成分が欠乏することにより疾患が発症するという新しい概念のはじまりであった.栄養素の欠乏は人体にとって重大な危機であり,その適応メカニズムに関してさまざまな角度から研究が進められている(第2章-8,9).
一方で,栄養素欠乏とは逆に栄養素の過剰摂取状態が長く続くことにより,内臓脂肪型肥満と糖尿病,脂質異常症,高血圧により動脈硬化のリスクが高まることは,現代社会の問題になっている.関連して,最近,分岐鎖アミノ酸代謝と肥満・糖尿病に関して新たな発見が報告されている(第3章-5,6).さらに,細胞内脂質代謝の異常により慢性炎症性疾患(代謝性疾患,がん,自己免疫疾患)のリスクが生じる(第3章-9).また,エネルギー代謝に重要なミトコンドリアの機能不全は,敗血症による複数の臓器システムの機能不全に影響することが示されている(第3章-8).一方,生活習慣病予防改善には適度な運動が有用であり,食事(脂肪酸)と運動による生体の変化(骨格筋機能)に関して解析がなされている(第2章-10).このような疾患の発症予防や未病状態の改善のために,個人ごとのビッグデータの解析・利用が重要であり「プレシジョン栄養」という新たな取り組みがなされている(第3章-10).
3)機能性食品について
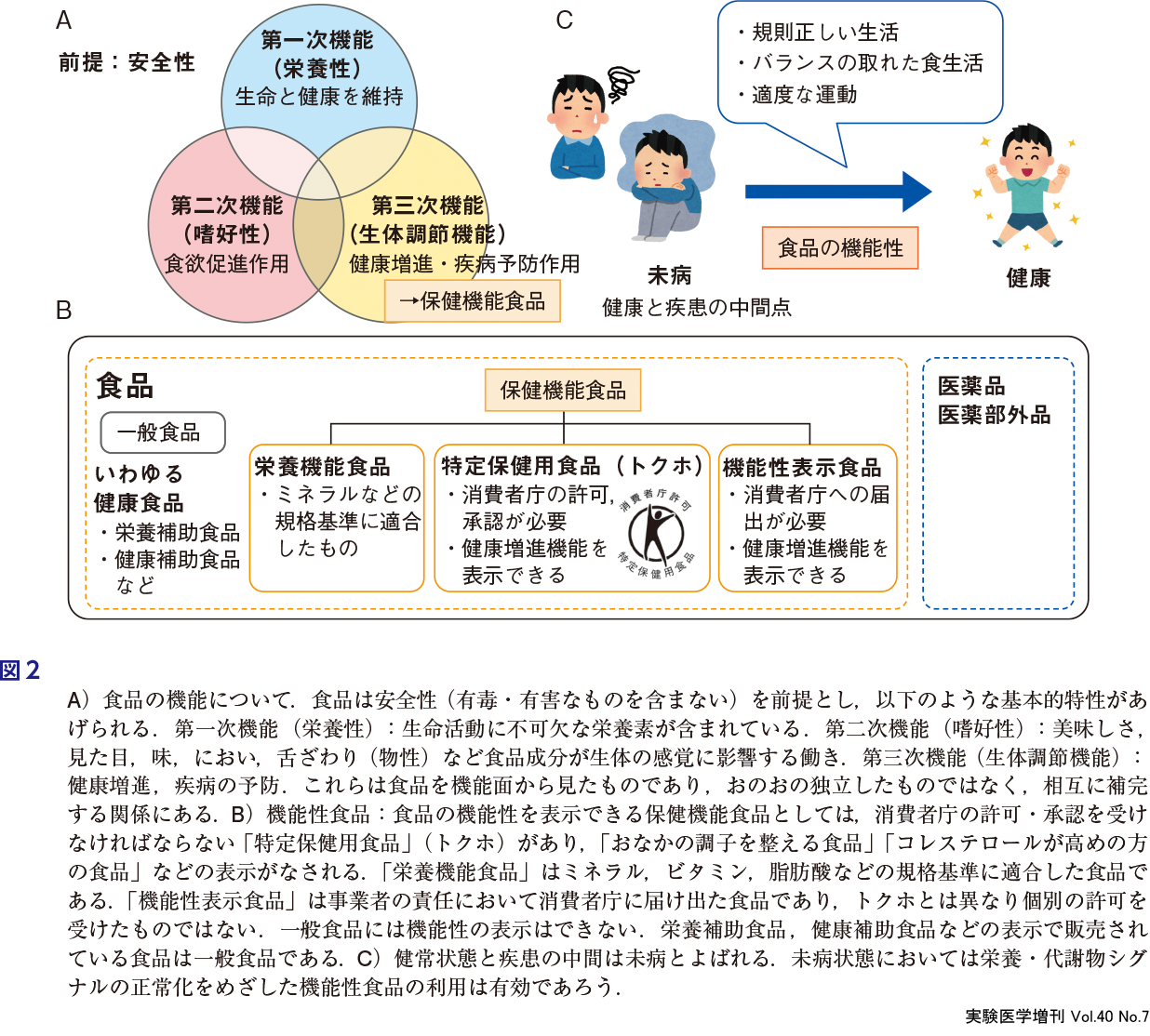
栄養学の発展のなかで,比較的近年に食品の機能性が注目されるようになった.食品の特性には,安全性を前提として栄養性,嗜好性,生体調節機能(生理機能性)などがあげられる(図2A).安全性は,毒物などの有害物質を含まない,腐敗・変敗していないなど食品としての必須条件である.栄養性とは五大栄養素などを含み,ヒトの生命と健康を維持する作用をもつことである.嗜好性とは味覚,嗅覚などを通じて食欲を促す作用を有することである.生体調節機能とは,ヒトの恒常性維持と,健康増進・疾病の予防など生体調節にかかわる機能である.1980年代には,農芸化学分野の研究者が食品の機能に関する研究を展開し,病気の予防効果をもつ食品を「機能性食品(functional foods)」と名付けた3).これがもとになり,食品に機能表示を可能にした特定保健用食品や機能性表示食品が市販されている(図2B).食品の特性を理解し,消費者自らが判断して選択・摂取することでバランスのとれた食生活を送ることができるような情報提供の手段として一定の有用性が認められる.この機能性表示食品の開発に関して最近ゲノム編集技術の利用が成功している(第1章-10).
健康と疾患の中間点の未病といわれる段階においては,(薬ではなく)食品の機能性が有用であると考えられる(図2C)4).古くから中国では「薬食同源」といわれているように,薬と食は同根でありバランスのとれた食事をとると健康が保たれるという考え方がある.逆に,薬と食物が相互作用することによって,予想外の作用をすることがあり注意が必要である.例えば,グレープフルーツが小腸での薬物吸収を促進することにより薬物の副反応が出るなど食品と薬物の相互作用が知られる(第1章-8).
2.食品機能の作用機序
1)核内受容体
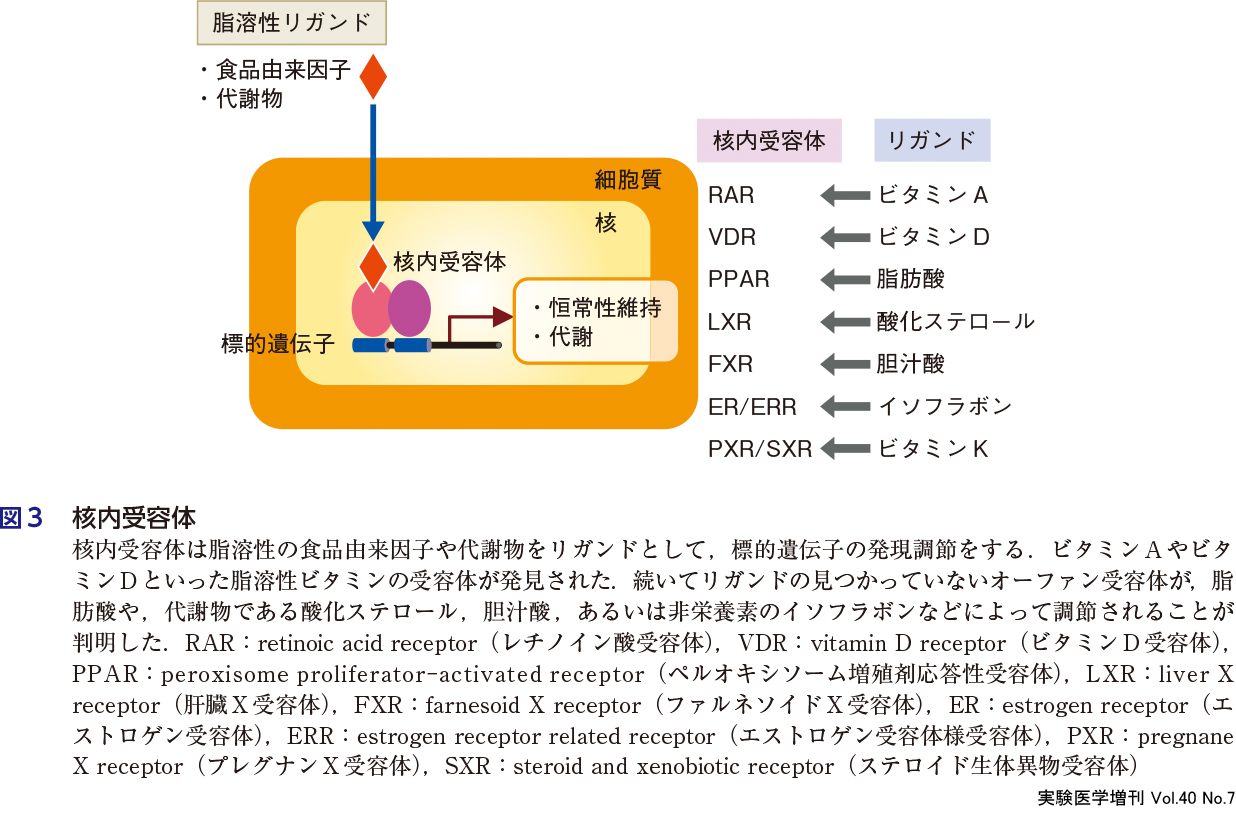
食品機能の作用機序が解明された最も重要な標的の1つに核内受容体(図3)の発見があげられるだろう.1980年代,核内ホルモン受容体がクローニングされ,リガンド誘導性の転写調節因子であることが判明した.すなわちステロイドホルモン類がその特異的な核内受容体を介して遺伝子の発現制御を転写レベルで行うことが明らかとなった5).グルココルチコイドやエストロゲンの受容体に引き続き,甲状腺ホルモンや脂溶性ビタミンであるビタミンDの受容体がクローニングされた.これらの受容体のアミノ酸配列は,特にDNA結合領域(Znフィンガー)において相同性が高く,遺伝子スーパーファミリーを形成していることが判明した.この相同性を利用して分子生物学的手法により未知オーファン受容体のcDNAのクローニングが可能となった.その成功例がビタミンA受容体(RARα)である6).cDNAのクローニング後にその転写促進能を手がかりにビタミンA受容体であることが明らかにされた.分子作用メカニズムが不明であったビタミンAの核内受容体が発見されたことで,ビタミンA研究が大きく発展した(第1章-5).
このような食品由来の微量成分が直接遺伝子発現制御をするという現象は生物学に大きなインパクトを与えた.その後,PPARやLXR,FXRといったオーファン受容体が発見されリガンドが同定されていった7).PPARはペルオキシソーム誘導剤であるクロフィブレートによって活性化されることが知られていたが,その後,脂肪酸によっても活性化されることが示された.LXRは酸化ステロール,FXRは胆汁酸といった食品由来の代謝物がリガンドになることが判明し,生体内の脂質代謝に重要な役割を果たすことが明らかにされた(図3).さらに,ビタミンKが核内受容体PXR/SXRのリガンドとして働き,胆汁酸代謝に影響することが報告された(第1章-6).
一方,核内受容体の転写活性化のメカニズムとして転写共役因子とのリガンド依存的な相互作用が重要であることが判明した8).転写共役因子にはアセチル化活性を有するものが存在し,後述のタンパク質修飾への重要性が示唆された.
2)代謝物によるシグナル修飾,エピジェネティクス
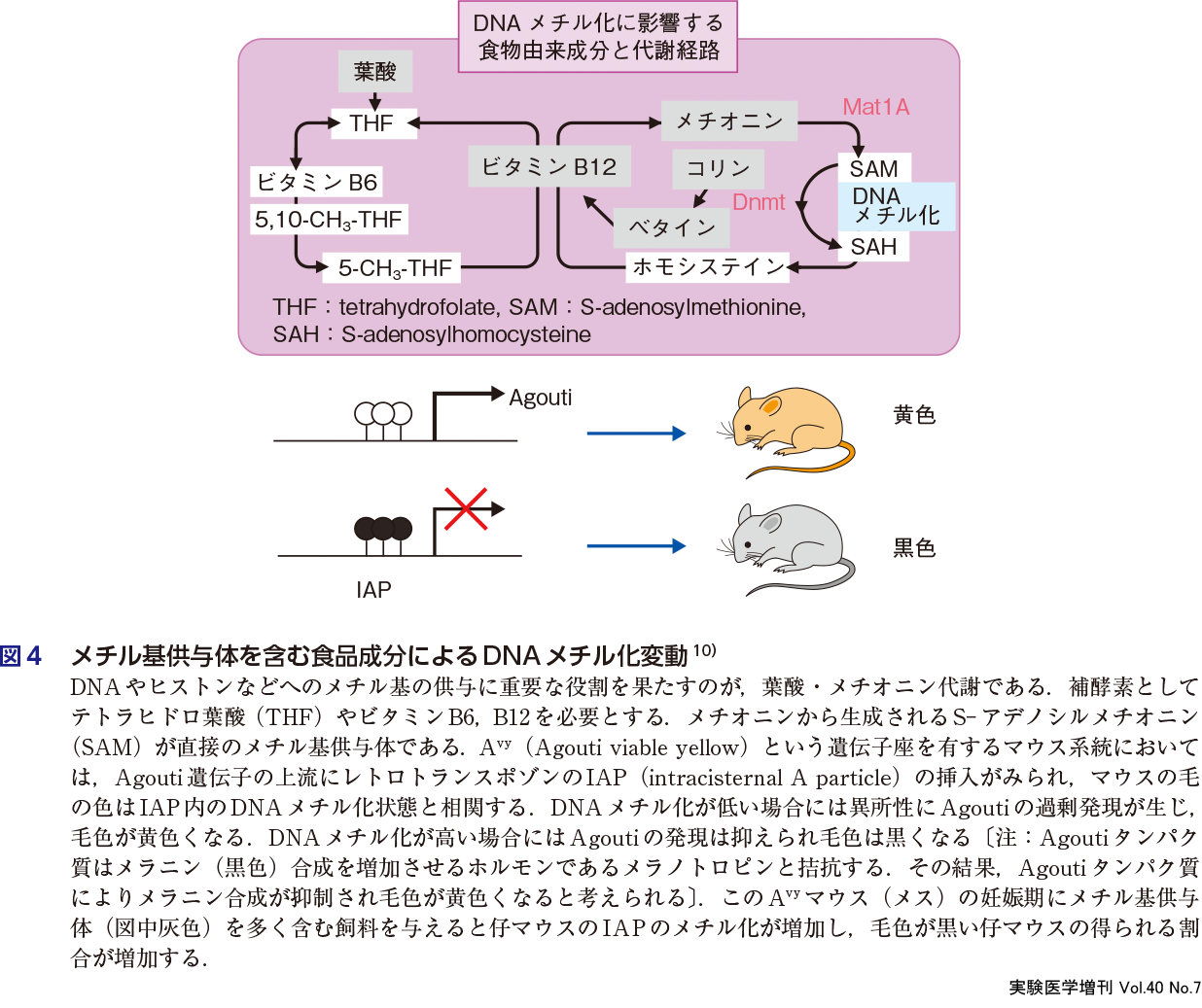
栄養素から生じるさまざまな代謝物は単なる中間産物とみなされてきた.しかし,栄養素代謝物はシグナル伝達,転写・エピゲノム制御など多彩な機能をもつことが明らかになってきた.代謝物はシグナル伝達物質としてエピゲノム修飾などに機能する場合がある.例えばグルコース代謝や脂肪酸代謝によって産生されるアセチルCoAがアセチル基の供与体としてヒストンや転写因子をタンパク質修飾(アセチル化)し,細胞増殖や分化にかかわる遺伝子の活性化を引き起こす9).またDNAやヒストンのメチル化反応に使われるメチル基供与体であるS-アデノシルメチオニン(SAM)はメチオニンや葉酸などさまざまな栄養素代謝を介して合成される.興味深いことに,メチオニン・葉酸・コリン・ベタイン・ビタミンB12などメチル基代謝に重要な食品成分を多く含む食餌をAvyという系統のマウス(妊娠期・母マウス)に与えると,仔マウスのDNAメチル化とそれに由来する表現型が変化しうることが報告された(図4)10).食品由来因子がエピゲノムに影響を与える例である.また胎児,乳児期,新生児期の栄養環境(栄養素の量や種類)は成長後の生活習慣病のなりやすさに影響し,DNAメチル化などのエピゲノムの関与が示唆される(第3章-1).実際,DNAメチル化は体の大きさと関連する.例えばハチはローヤルゼリー(≒DNAメチル化阻害)摂取により女王蜂になる.今後の解析に有用な技術として,最近,エピゲノム編集によりDNAメチル化の操作が可能となった(第3章-2).一方,DNA以外にもヒストンや遺伝子発現に関連するタンパク質のリジン残基やアルギニン残基が,翻訳後修飾としてメチル化を受けることが知られる.またヒスチジンのメチル化が最近注目されている.メチルアミノ酸はeNOS阻害による血圧上昇などの生理作用や,筋分解の血中マーカーなどとして利用される(第3章-3).
加えて,αケトグルタル酸などのTCA回路の中間代謝物はヒストン脱アセチル化酵素や,メチル化シトシン水酸化酵素(DNA脱メチル化酵素,TET)の補酵素としてエピゲノム修飾に働く11).酪酸やβヒドロキシ酪酸(ケトン体)はヒストン脱アセチル化酵素の阻害剤として働く(ヒストンのアセチル化を増やす)12).さらに食餌由来のリボフラビン(ビタミンB2)から合成されるFADはLSD1(リジン特異的脱メチル化酵素1)などの補酵素として働き,エネルギー代謝や細胞機能を調節する(第3章-4).同じくビタミンB群から合成されるNAD+はSirt1(脱アセチル化酵素)の補酵素として働く.NAD+/NADHの比が上昇する状態,すなわち低栄養の状態でSirt1は活性化され,ヒストンを含めた基質タンパク質の脱アセチル化を行う.また細胞内のAMP/ATP量はAMPキナーゼ活性を制御し,cAMPはプロテインキナーゼを活性化する.このように細胞内の代謝物はさまざまなシグナル伝達物質として働くことが知られている11).
3)腸内細菌の関与と腸を起点とした臓器連関
ⅰ)食品成分と腸内細菌叢:腸管と臓器連関
食品の代謝変化と機能性を論じるにあたり,腸内細菌叢(gut microbiota)が重要な役割を果たすことが注目されている.すなわち消化器官(小腸や大腸)の管腔内(トポロジー的に体の外側)に存在する腸内細菌叢の生成する短鎖脂肪酸などの代謝物がシグナル伝達に果たす役割が明らかにされてきた.ヒトの腸管内には,数百種類以上,100兆個にも及ぶ腸内細菌が生息しているとされる.腸内細菌の集団を腸内細菌叢とよぶが,これらは宿主であるヒトが摂取した食物によってその構成が変化する.また腸内細菌叢の構成や,腸内細菌が発酵などにより産生する代謝物が,腸管(上皮細胞や免疫細胞)に作用し,腸内環境を含む生体機能全体に影響を与えると考えられている(図5).例えば,腸内細菌が産生する短鎖脂肪酸はGタンパク質共役型受容体を介して免疫系を活性化することが知られている(第2章-2)13).また腸内細菌で代謝された脂肪酸代謝物が褐色脂肪組織のエネルギー代謝を調節する(第1章-2).さらにフルクトースは腸内細菌叢で短鎖脂肪酸になり,肝臓での脂肪酸合成に利用され脂肪肝形成の一因となることが報告されている(第3章-7).以前は,食物繊維などの難消化性多糖類はヒトが消化・利用することのできない不要な栄養素であると捉えられていたが,近年の腸内細菌研究により,腸内細菌に資化されることによって産生された短鎖脂肪酸が,宿主のエネルギー源としてだけでなく,生体恒常性維持に重要な役割を果たすことが明らかにされている.
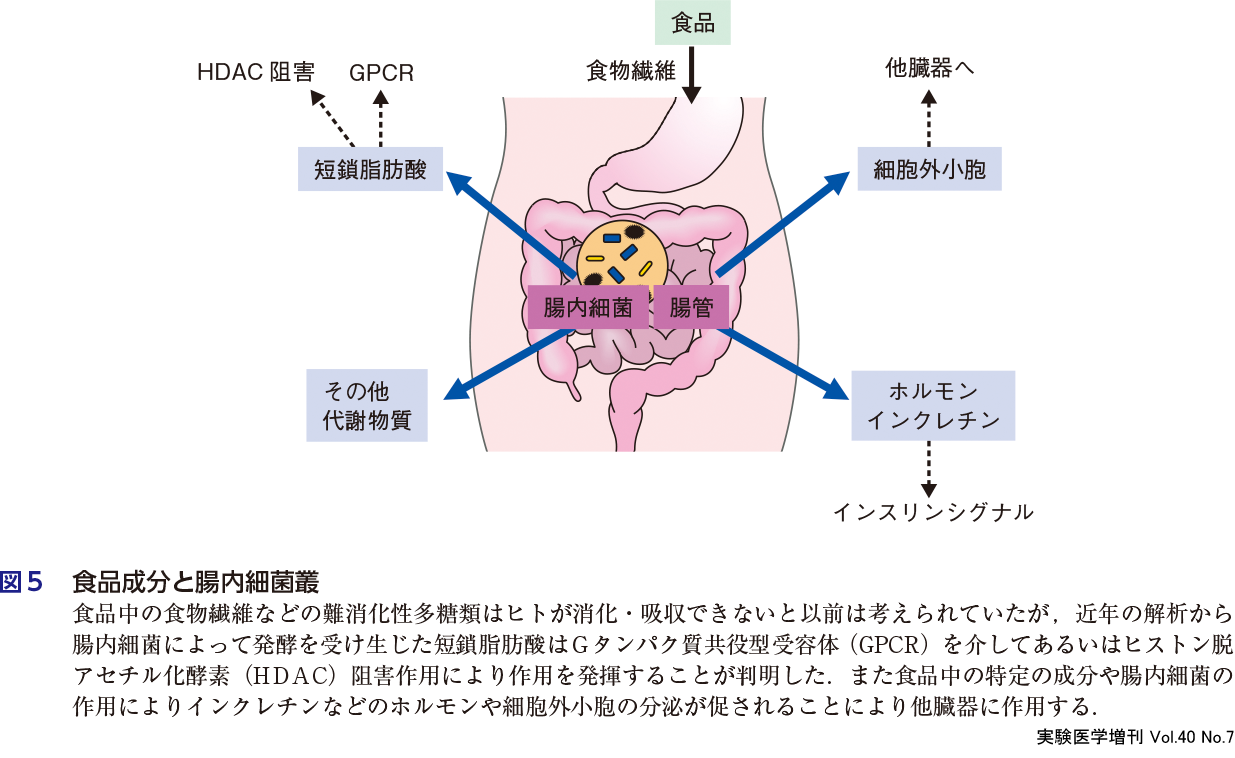
ⅱ)他臓器との臓器連関
このように腸管を出発点として他臓器との臓器連関が知られている.食品成分による臓器間の相互作用として細胞外小胞(extracellular vesicle, エクソソーム:細胞から分泌される膜小胞)が注目される.例えばGABAやカルノシン(ジペプチド)によって腸管から細胞外小胞分泌が促進され,細胞外小胞中のmiRNAが神経細胞を活性化する(第2章-4)とともに,食品中に含まれる細胞外小胞中の高分子(miRNAなど)が食品を摂取したホスト(ヒト)に作用する可能性が示されている(第1章-8).また,腸管内分泌細胞の管腔側には食物の分解産物に対する受容体が存在し,消化管ホルモン(例,インクレチン)を分泌,神経を介して脳に作用し食欲などに影響する(第2章-5).ショウジョウバエモデルでは,腸内細菌由来の酢酸がインスリンシグナルや個体の成長を保つことが示されている.すなわち摂食依存的に腸内分泌細胞からNRFという分子(≒インクレチン)が放出され,インスリン様シグナルを活性化する(末梢の同化シグナルが活性化する)(第2章-3).そして腸内細菌は肥満や糖尿病などの代謝疾患の発症に影響することが知られている14).近年さまざまな研究知見と技術の進歩によって,腸内細菌叢を遺伝子レベルや代謝物レベルでの網羅的解析により可視化できるようになり,食と健康長寿の相関を科学的に明らかにする試みが進んでいる.経験によって語られてきた食と健康の関連を,分子レベルで記述することが可能になってきた.
一方,腸管以外でも栄養・代謝物シグナルと臓器連関に関して研究が進んでいる.NAD+がSirt1を活性化することは前述したが,NAD+合成中間体のNMNの摂取は抗老化作用を示す.脂肪組織からNAMPT(NMN合成酵素)が細胞外小胞として分泌され他臓器でNAD+を増加させ抗老化作用を示すことが明らかになっている(第2章-7).他方,嗅覚受容体が膵臓β細胞で発現しオクタン酸(中鎖脂肪酸)を認識しインスリン分泌を促すことが示された(第2章-6).また分岐鎖アミノ酸であるバリンの代謝物であるβアミノイソ酪酸(BAIBA)は運動時に骨格筋から分泌され,白色脂肪細胞のベージュ化を引き起こし,エネルギー代謝を活性化する15)ほか,動脈硬化の進展を抑制することが知られる16).すなわち臓器間の情報伝達にも代謝物が利用されていることが明らかになっている.
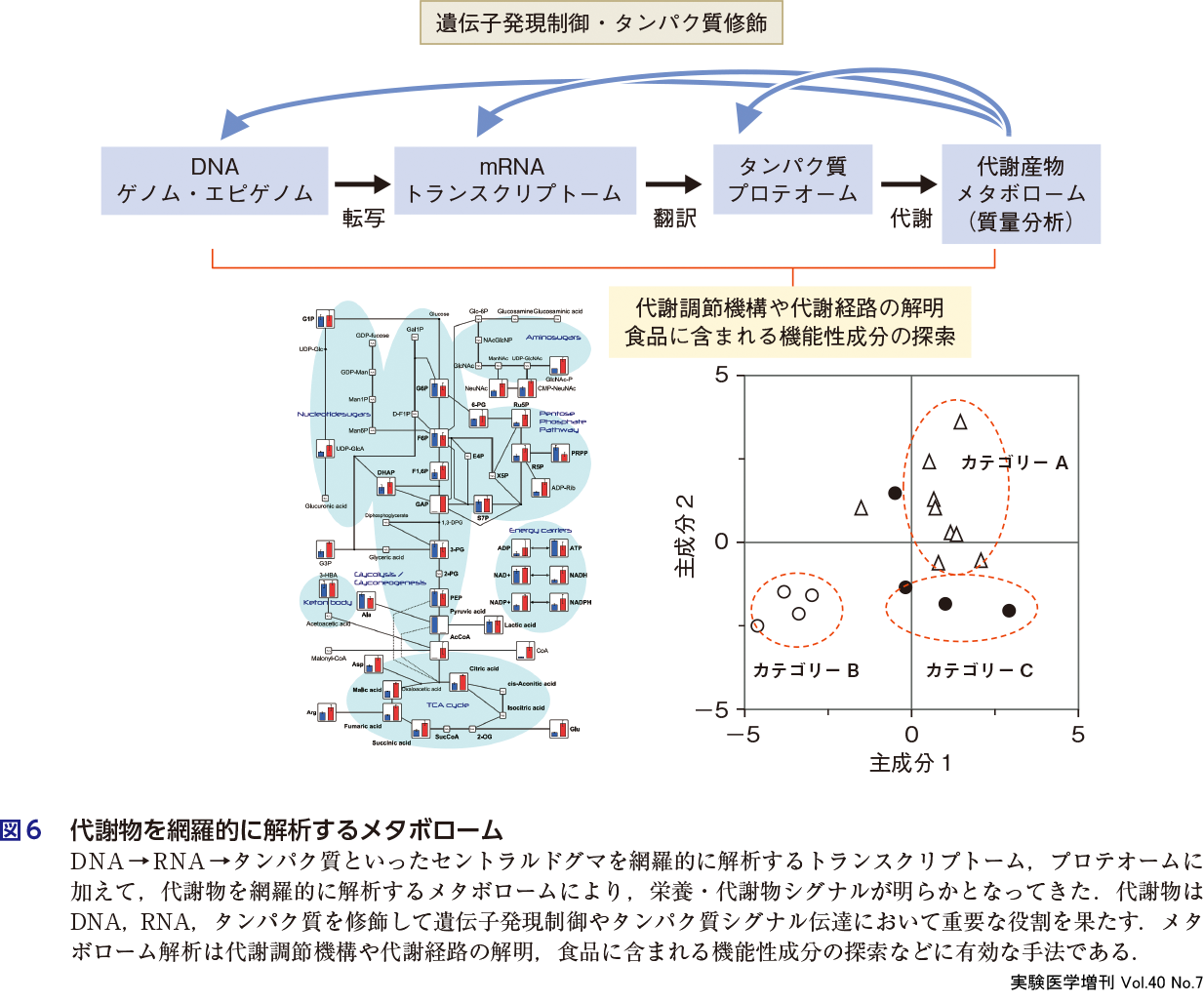
4)メタボローム
近年の代謝研究の急速な進展は代謝物測定技術の向上によるところが大きい.質量分析によりどのような代謝物が存在するか,どのような修飾を受けたのか正確に理解することが可能となってきた17).トランスクリプトームやメタボロームは生体内の多数の構成成分の変化を,バイアスをかけない網羅的な手法によって解析し,生命現象を包括的に理解しようとするものである(図6).栄養・代謝物シグナルの進展にメタボロームが貢献している.種々の質量分析計などの技術革新によりメタボローム測定法が開発され,遺伝子情報の解析結果と組合わされ,代謝調節機構の解明や食品に含まれる機能性成分の探索など多様な応用がなされている.代謝経路に関してはデータベース(例,KEGG)の構築により,代謝物と代謝物の関係,代謝物と代謝酵素との関係が明らかとなってきた.さらに,生体内の代謝のみならず,食品および食品に含まれる機能性成分の評価についてメタボロームの手法が活用されている(第1章-9).
おわりに
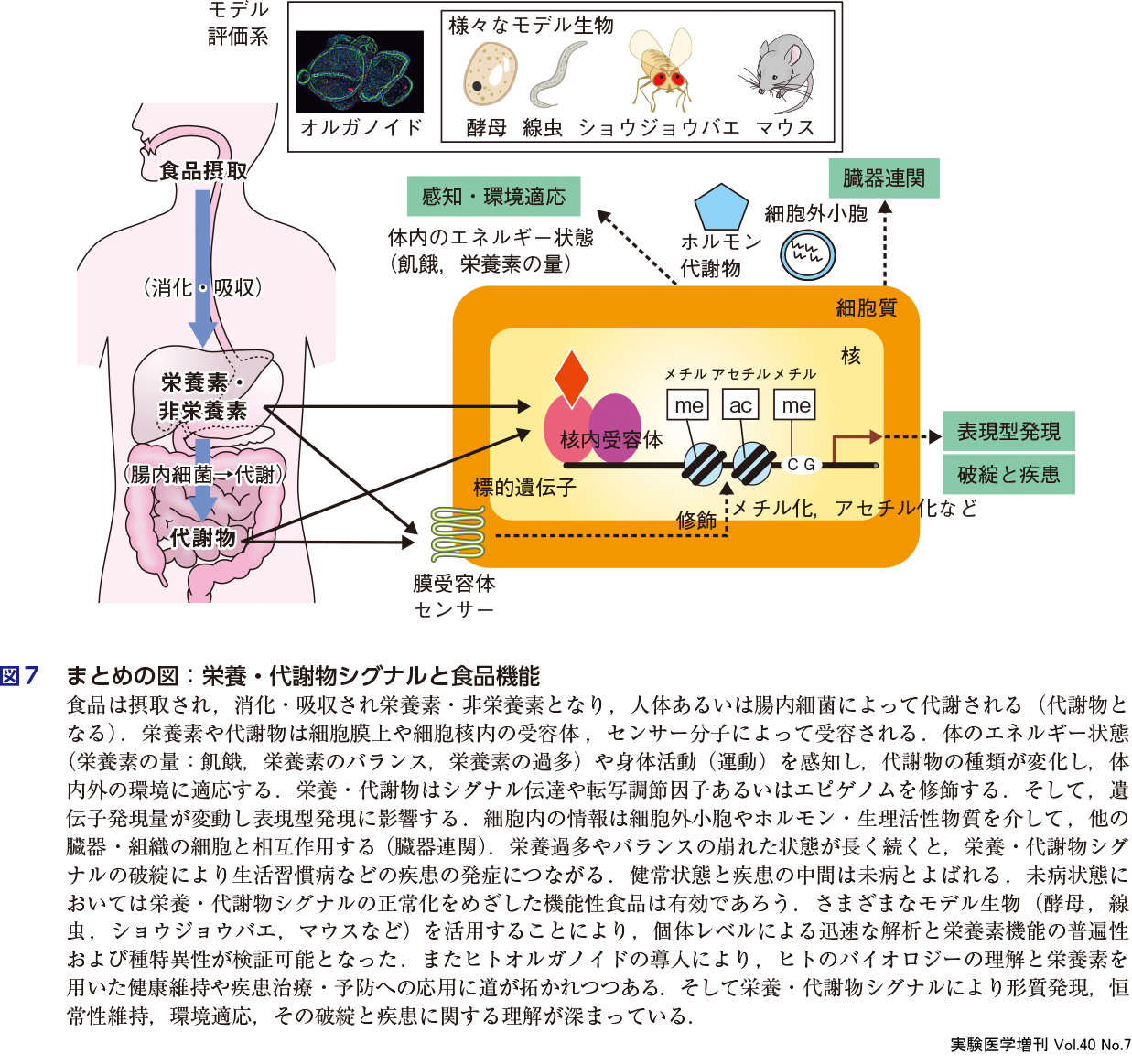
これまでの研究により,食物の消化・吸収・排泄などの動態,栄養素の代謝経路・生化学的特性が明らかにされてきた.栄養学研究と代謝物研究は,以前は異なる研究領域で実施されていた.栄養学研究はヒトや動物個体における豊富な知見を有し,特に栄養素の消化・吸収・輸送・代謝に関する研究が行われてきた.代謝物研究は分子生物学やメタボロームなどの先端技術を用いた解析を得意とし,栄養素がどのようにして生命情報に変換されるのかという栄養素の感知,シグナル伝達,転写調節に関する研究が盛んであった.近年,両者(栄養学研究と代謝物研究)の協同により,生体における栄養素の流れ(摂取・感知から機能発現までの一連の流れとそのしくみ)が捉えられるようになった.具体的には栄養学,生化学,発生学,分子生物学,代謝生理学など複数の分野に跨る研究により,さまざまなモデル生物(酵母,線虫,ショウジョウバエ,マウスなど)を活用することで,個体レベルによる迅速な解析と栄養素機能の普遍性および種特異性が検証可能となった.またiPS細胞を用いたヒトオルガノイドの導入により,ヒトのバイオロジーの理解と栄養素を用いた健康維持や疾患治療・予防への応用に道が拓かれつつある.例えば,小腸オルガノイドが構築され,栄養素や食品成分の吸収・代謝・機能をより正確に評価できるようになった(第2章-1,図7).このように栄養素の機能が組織・細胞レベルで明らかにされつつある.特に,栄養素が細胞によってどのように感知され,シグナル伝達や転写・エピゲノム形成に寄与するかが明らかになってきた.またその結果,高次生命現象(発生,疾患,老化)に帰結するかに関して研究がなされている.
本増刊号では,「第1章,食品の機能性と栄養・代謝物シグナル」「第2章,栄養・代謝物シグナルと臓器連関」「第3章,栄養・代謝物シグナル修飾および破綻と疾患」というように章立てしている.これらの各章は緩やかな分類であり相互に密接にかかわりのあるものである.本増刊号は栄養・代謝物シグナルと食品機能の最前線の紹介であり,今後の研究の発展・議論の端緒となることを期待したい.
文献
- 「生物と無生物のあいだ」(福岡伸一/著),講談社,2007
- 「楽しくわかる栄養学」(中村丁次/著),羊土社,2020
- Swinbanks D & O’Brien J:Nature, 364:180, doi:10.1038/364180a0(1993)
- 日本学術振興会 産学協力委員会 R021食と未病マーカー委員会
- Chambon P:Mol Endocrinol, 19:1418-1428, doi:10.1210/me.2005-0125(2005)
- Giguere V, et al:Nature, 330:624-629, doi:10.1038/330624a0(1987)
- 梅園和彦:オーファンレセプターの機能解析.実験医学,16:2431-2435, 1998
- Kamei Y, et al:Cell, 85:403-414, doi:10.1016/s0092-8674(00)81118-6(1996)
- Ogryzko VV, et al:Cell, 87:953-959, doi:10.1016/s0092-8674(00)82001-2(1996)
- Waterland RA & Jirtle RL:Mol Cell Biol, 23:5293-5300, doi:10.1128/MCB.23.15.5293-5300.2003(2003)
- Sharma U & Rando OJ:Cell Metab, 25:544-558, doi:10.1016/j.cmet.2017.02.003(2017)
- Shimazu T, et al:Science, 339:211-214, doi:10.1126/science.1227166(2013)
- Lee WJ & Hase K:Nat Chem Biol, 10:416-424, doi:10.1038/nchembio.1535(2014)
- Cho I, et al:Nature, 488:621-626, doi:10.1038/nature11400(2012)
- Roberts LD, et al:Cell Metab, 19:96-108, doi:10.1016/j.cmet.2013.12.003(2014)
- Shimba Y, et al:Biol Pharm Bull, 43:1016-1019, doi:10.1248/bpb.b20-00078(2020)
- Soga T, et al:J Proteome Res, 2:488-494, doi:10.1021/pr034020m(2003)
著者プロフィール
亀井康富:京都大学農学部食品工学科卒業(農学博士),カリフォルニア大学,大阪バイオサイエンス研究所,国立健康・栄養研究所,東京医科歯科大学を経て,2012年より京都府立大学大学院生命環境科学研究科分子栄養学研究室教授.栄養・代謝物シグナルと食品機能について骨格筋を中心に研究しています.