
概 論
セントラルドグマにまつわる32の新常識から知る生命科学のトレンド
田口英樹1),小林武彦2),稲田利文3)
(東京工業大学科学技術創成研究院細胞制御工学研究センター1),東京大学定量生命科学研究所2),東京大学医科学研究所3))
1第1章 DNAの新常識
「遺伝子は解読から創生へ,DNA複製は高次元レベルの解析へ」
セントラルドグマは遺伝子の発現スキームであり,その最初にあるのが遺伝子DNAとその複製になる.
筆者(小林)が大学院生だった30年前,DNA複製の研究は分子生物学の中心の1つであった.大腸菌のプラスミドの複製開始機構を筆頭に,大腸菌ゲノムの複製機構,さらに真核細胞では出芽酵母の複製開始点がクローニングされ,そこで働く複製開始因子,細胞周期との連動など,次々に新しい発見があった.ポリメラーゼや変異・修復のメカニズムも続々と解明され,まさに分子生物学の絶頂期とも思える頃であった.ところが哺乳動物の複製開始点については壁にぶつかった.まずサイズが大きすぎて当時のクローニング中心の解析方法ではどうにもならなかった.さらに,決まった配列から開始反応がはじまるわけではなく,当時まだよくわからなかったクロマチン構造などの影響が大きいと考えられた.転機はゲノムプロジェクトとともに現れた.DNAチップや次世代シークエンサー(NGS)を使ったハイスループット解析により,複製開始や進行状況などが包括的に調べられるようになった.
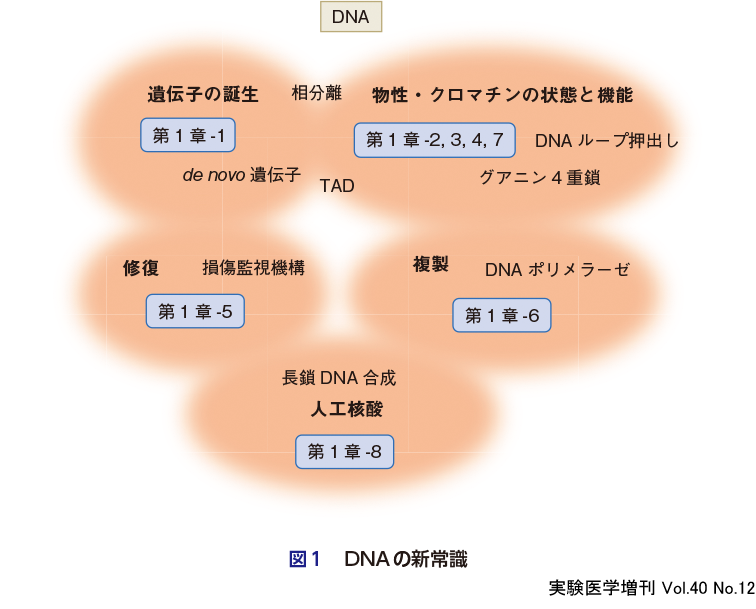
本章ではDNAの複製を中心に,10名の研究者による常識を打ち破る研究を紹介する(図1).
山内・岩崎は,セントラルドグマのスタート地点である遺伝子がそもそもどのようにできたのか,その謎に迫る.これまでよく知られていた遺伝子増幅ではない「読み枠のずれ」などによる新規の遺伝子の創出について提案している(第1章-1).末次は大腸菌のin vitro DNA複製系を利用し,長鎖DNAの合成システムの構築に成功した(第1章-8).平谷はヒト細胞の複製開始のタイミング,クロマチン構造との関係,開始反応の空間的制御を詳細に解析している(第1章-4).大学はNGSを用いて,ヒト細胞の複製フォークのリーディング鎖とラギング鎖を区別することで開始点のマッピング,進行状況のモニターなどのゲノムワイドな解析を行っている(第1章-6).坂田・西山は複製後の姉妹染色分体をつなぐコヒーシンが,DNAを押し出し,プロモーターとエンハンサーを近づけて転写を制御するという新機能について,イメージングを中心とした研究を行っている(第1章-2).また,島添らは,クロマチンの物性について,これまでは直径30 nmの「硬い」構造体と思われていたが,じつは細く柔らかな流動性のある物質であることを発見した(第1章-3).日下部・菅澤はこれまで謎であった修復タンパク質がいかにDNAの傷を見つけるのかについて,修復タンパク質は自由拡散によりランダムにDNAに結合し,その滞留時間がDNAに傷がある場合は長くなり修復反応につながることを提唱している(第1章-5).正井は,DNA自身がつくる構造体「G4構造」がゲノム中に広く存在し,複製などさまざまな機能をもっていることを紹介している(第1章-7).このように遺伝子は読む時代からつくる時代へ,DNAの複製は配列の研究から高次元の構造と時空制御の解析へとシフトしている.
2第2章 転写の新常識
「新・RNAワールド」(図2)
セントラルドグマにおける「転写」は,遺伝子のコピーであるmRNAの生産が主となり,タンパク質へとつなぐ媒介的な役割として考えられていた.一方,生命の誕生のプロセスでは,最初にRNAが登場し(RNAワールド),おそらく次にはリボソームができてタンパク質がつくられ,最後に遺伝情報のストックとしてDNAが登場したと推察されている.つまり,登場した順番から考えるとDNAの方が「RNAのコピー」と捉えることもできる.リボソームRNA(rRNA)やトランスファーRNA(tRNA)など,RNA自身が触媒作用をもつものでは,両者の関係がよりはっきりと見てとれる(小林,第2章-6参照).
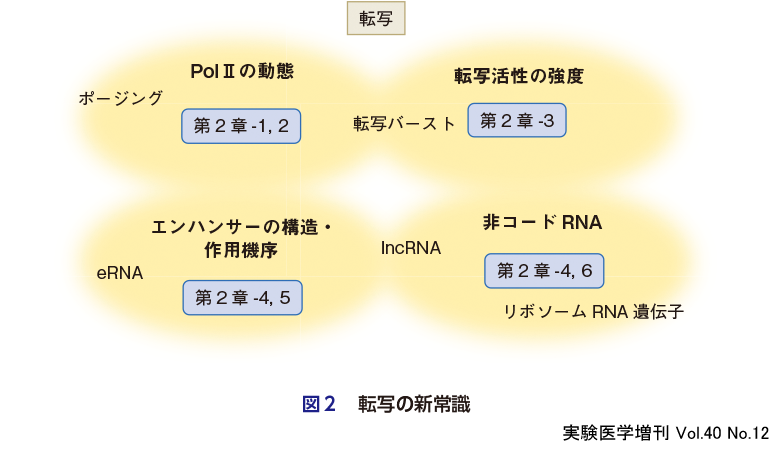
鯨井・胡桃坂は長年の謎であるRNAポリメラーゼⅡ(PolⅡ)のヌクレオソーム通過時のふるまいをクライオ電子顕微鏡で詳細に観察した.その結果PolⅡはヌクレオソームを剥がしながら進み,転写伸長因子は「剥がす反応」を助けることがわかった(第2章-1).高橋は,その転写伸長因子には,PolⅡの一時的な停止(ポージング)を解除する役割があることを紹介している.これまではPolⅡのロード量のみが転写活性を左右すると考えられていたが,転写のポージング解除も重要な調整要因になっている(第2章-2).またエンハンサーの働きについても重要な進展があった.深谷は1細胞レベルの遺伝子発現の不均一性をイメージングで捉え,数分単位でON/OFFをくり返す現象「転写バースト」を観察している.転写の強さはこのバーストの頻度によって決まっていることが示唆されている.また,これまでの常識を覆す1つのエンハンサーが複数の遺伝子に影響を与える「ハブ」機能も見出している(第2章-3).堀江らはホヤの解析から,エンハンサーに結合する複数の転写因子の位置関係について新しい法則を発見した.つまり転写因子間の距離が近いと発現のタイミングが早くなる(第2章-5).斉藤らはエンハンサーの作用機序として,そこからの非コードの転写活性(エンハンサーRNA,eRNA)を見出している.eRNAはエンハンサーとプロモーターをつなぐ役割を担い,また液-液相分離(LLPS)を促しクロマチンの塊であるTAD(topologically associating domain)の形成を介して複数の遺伝子を活性化する「スーパーエンハンサー」構築にも関わる.これらの因子は,新たな抗がんのターゲットとしても期待される(第2章-4).最後の私たち(小林)の研究は,出芽酵母の寿命を決める非コードの転写活性を見出した.その転写が走るとコヒーシンの結合状態など,修復反応に影響をおよぼしゲノムの安定性を低下させて老化を誘導する(第2章-6).
以上のように転写反応の開始,伸長の動的なふるまいにおける新常識に加えて,非コードRNAが生命現象を司る重要な鍵になってきた.
3第3章 RNAの新常識
「変幻自在なRNA機能の発見」
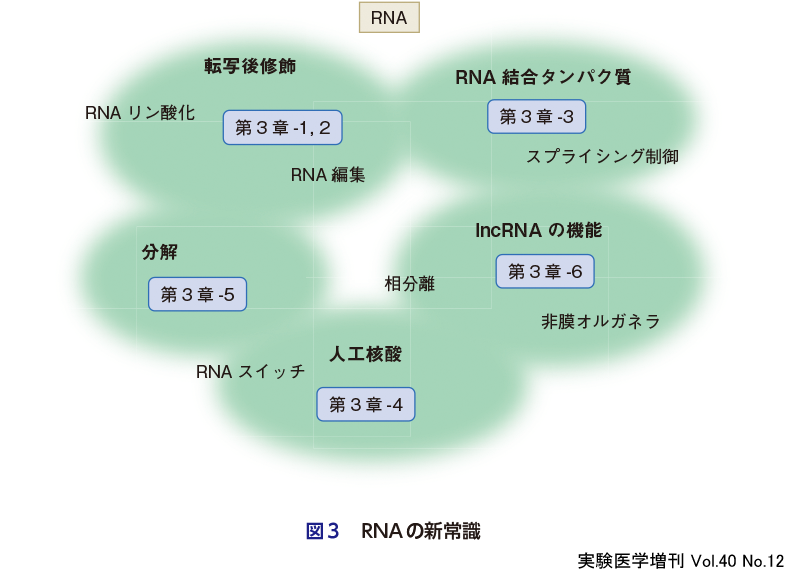
遺伝子発現解析手法の発展により,高感度かつ高精度での発現情報が明らかになり,RNAの常識が大きく変貌しつつある(図3).遺伝子発現の重要な制御因子としてのRNA修飾の多彩な機能が次々と解明され,未開拓の領域であったRNAエピトランスクリプトームの世界が大きく広がろうとしている.mRNAワクチン開発の基盤となるRNA修飾の免疫誘導機能や,m6A修飾によるmRNA機能制御機構の解明が契機となり,RNA修飾の同定と新規機能の解明が進み,現在150種類以上のRNA修飾が発見されている.tRNAのコドン解読におけるRNA修飾の新規機能を解明してきた鈴木らの研究グループが,tRNA修飾解明の経緯とtRNAリン酸化の新規機能を紹介する(第3章-1).ゲノムに書き込まれた遺伝情報は,転写後修飾の一種であるRNA編集によって書き換え,または書き足し,を受ける.脱アミノ化修飾は,コドン置換やmicroRNAの標的mRNAの認識などに影響する.さらに外因性RNAと識別できる目印として自己RNAに付加され,その破綻と自己免疫疾患の関連が解明されつつある(河原,第3章-2).RNA編集による遺伝性疾患の治療をめざす動きも今後の進展が期待される.
RNAの機能の多くは,RNA結合タンパク質(RBP)と複合体を形成して発揮される.RBPは,セントラルドグマのさまざまな段階での制御機構に必須な機能を果たす(武内,第3章-3).RBPはmRNAと複合体を形成し,膜構造をもたない細胞内オルガネラである液-液相分離(LLPS)による液滴形成を促進し,神経変性疾患の主因としても注目されている.真核生物ゲノムの大部分を占める非コード領域から数万種類もの長鎖非コード(long non-coding,lnc)RNAが産生されるが,その多くは機能未知である.なかでも,細胞内構造体の骨格として働くものはarchitectural RNA(arcRNA)とよばれ,さまざまなRBPが集合体を形成して細胞内相分離を誘発し,巨大な非膜オルガネラを形成する.arcRNA依存的に形成される非膜オルガネラの構造と形成原理について第3章-6(廣瀬ら)で概説する.
mRNAでは合成のみでなく「分解」も積極的な意義をもっている.例えば,免疫応答や炎症にかかわる遺伝子の発現の異常は,免疫システムの暴走を招くが,RNA分解酵素であるRegnase-1は,免疫応答や炎症関連mRNAの適切な分解を担うことで,免疫システムの適正な作動を支えている(赤木・竹内,第3章-5).
遺伝子発現制御を担うRNAエレメントとして,化学反応触媒をもつリボザイム,リガンドとの相互作用により転写や翻訳を調節するリボスイッチなどが機能している.RNAエレメントを組合わせることで,遺伝子発現を制御可能な人工RNA「RNAスイッチ」の開発が進んでいる(小野・齊藤,第3章-4).さまざまな細胞内タンパク質やRNAの存在状態に応じて目的の遺伝子発現を自由に制御できる日も遠くないかもしれない.
4第4章 翻訳の新常識
「非典型的な翻訳の拡がりと多彩なリボソームの機能」
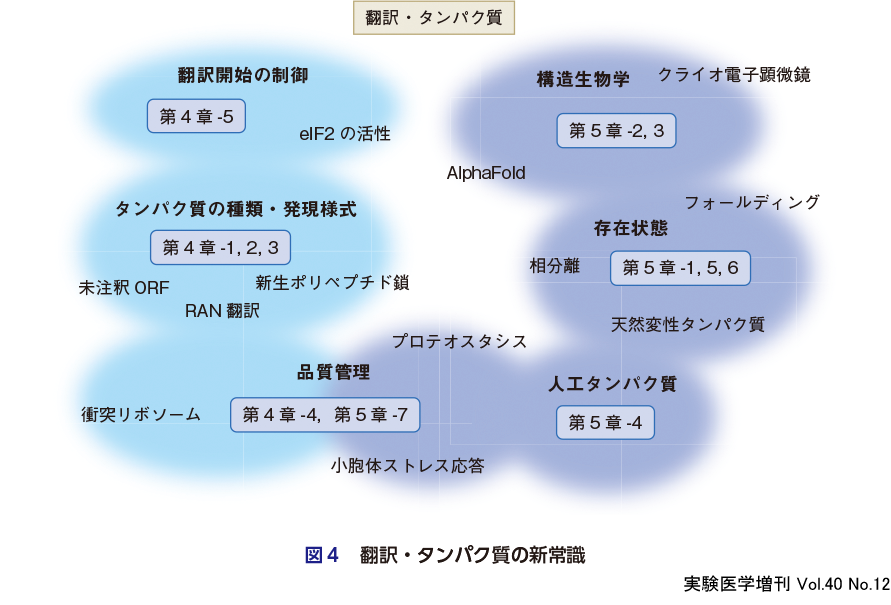
DNAにある遺伝情報がmRNAやtRNAを介してアミノ酸配列に変換されてタンパク質(ポリペプチド)が合成される翻訳はセントラルドグマの最終ステップである.基本的には,リボソームがmRNAの開始ATGコドン(mRNAではAUG)から翻訳を開始し,コドンにしたがってアミノ酸をつなげて,終止コドンで翻訳を終了する,というのが翻訳の概略である.しかし,この10年ほどの間に,翻訳の常識は大きく書き換わりつつある(図4).
まず,開始コドンはいつもATGでないことがわかってきた.開始コドンは(特に真核生物では)ほぼATGと従来考えられていたが,CTGなどの非ATGでの翻訳開始が思いの外多いことが明らかになっており,なかには疾患に関与する場合もある(浅野,第4章-1).また,細胞内の翻訳状況をNGSで精度高く網羅的に調べるリボソームプロファイリング法の発展により,ノンコーディングRNAの一部から翻訳が起こって,既知のタンパク質の世界を凌駕する未開拓のタンパク質の世界が拡がっていることが明らかになってきた(市原ら,第4章-3).さらに,翻訳の主役のリボソームの周辺でも常識が変わりつつある.リボソームはいつも同じ速度で翻訳伸長するわけではなく,頻繁に一時停止しうること,また,合成途上の新生タンパク質自身が「完成前」に生理機能を持ちうることもわかってきた(千葉・藤原,第4章-2).また,リボソームは,細胞内タンパク質の「誕生」の場であると同時に,うまく合成されない翻訳産物(タンパク質)を検知して「分解」する「品質管理」の場であることがわかってきた.つまり,リボソームは,「正常な」遺伝子産物を認定するセントラルドグマの最終管理者と位置付けられる(稲田,第4章-4).近年,多くの生命現象が翻訳レベルで制御されていることが明らかになりつつあるが,その例の1つに翻訳開始制御による細胞ストレスへの対処がある.この制御機構の立体構造基盤についても紹介する(柏木・伊藤,第4章-5).
5第5章 タンパク質の新常識─
「変容し拡大するタンパク質の世界」
細胞内のあらゆるタンパク質はリボソームを介した翻訳で産み出され,アミノ酸配列に従って立体構造を形成(フォールディング)して機能する,というのが古典的なタンパク質の常識である.しかし,ここ数年の間に従来のタンパク質像が大きく変わりつつある(図4).
まず,タンパク質の総種類が増えている.従来,細胞内のタンパク質の総体(プロテオーム)はDNA配列から予想される遺伝子読み枠(ORF)で定義されていたが,第4章の翻訳の新常識でいくつか紹介があったように,既知のORF以外から翻訳が起こって,タンパク質が多数できることがわかってきている.
タンパク質のフォールディングに関して,1950年代にはフォールディングは自発的に起こるというドグマ(いわゆるAnfinsenのドグマ)が確立したが,その後,細胞内ではシャペロンがフォールディングを助けていること,フォールディングが異常なタンパク質は凝集体となり,その一形態であるアミロイドやプリオンは深刻な疾患にかかわることがわかってきた(田口,第5章-1).ここで,細胞内でタンパク質がリボソームで誕生してから分解されるまでを「タンパク質の一生」とよぶことがあるが,最近では,より広い概念である細胞内のタンパク質の恒常性(プロテオスタシス)が提唱されている.特に,小胞体でのプロテオスタシス維持は重要である(上垣ら,第5章-7).
タンパク質の立体構造についても多くのブレイクスルーや革命が起こっている.タンパク質の立体構造を決定する構造生物学の圧倒的な主流はX線結晶構造解析であったが,この10年ほどの間にクライオ電子顕微鏡がとって代わろうとしている(稲葉・張,第5章-2).関連して,アミノ酸配列だけから計算科学でタンパク質の立体構造を予測するのは困難と思われていたが,2021年にAlphaFold2という構造予測の革命が起こった.この革命により,実験的に立体構造を解かなくてもかなりの高精度でタンパク質の立体構造を入手して研究に使える時代になった(木原,第5章-3).また,立体構造予測の発展に伴って,以前は不可能といわれていた生物がもたないタンパク質の合理的な設計(de novoデザインタンパク質)も可能となりつつある(古賀ら,第5章-4).さらにいえば,そもそも立体構造をとらないタンパク質がじつは多数あり,真核生物のタンパク質の3割もが立体構造をとりえない「天然変性タンパク質(天然変性領域)」であることがわかってきたのも常識を覆した1つである(太田ら,第5章-6).
最後に,細胞内でのタンパク質の存在状態にも新たな視点が加わった.タンパク質(に加えて核酸など細胞内因子も)は状況に応じて液-液相分離して液滴をつくることが生理的に重要であることがわかってきている(野田,第5章-5).
おわりに
以上,どの章にも,新常識,新知見,ブレイクスルー,革命,……といった用語が満載である.興味のあるところから読んでいただき,読者一人ひとりにセントラルドグマに関する新たな視点が加われば幸いである.
著者プロフィール
田口英樹:1989年東京工業大学工学部卒業.’93年東京工業大学大学院総合理工学研究科博士課程修了(吉田賢右教授).博士(理学).日本学術振興会特別研究員,東京工業大学資源化学研究所助手,東京大学新領域創成科学研究科准教授を経て,2010年より東京工業大学生命理工学研究科教授.’17年より東京工業大学科学技術創成研究院細胞制御工学研究センター教授.’02年〜’06年JSTさきがけ研究者を兼任.ちょうど30年前に大学院でたまたまはじめたシャペロン研究は当時のタンパク質科学の常識(Anfinsenのドグマ)への挑戦であり,新しいタンパク質研究を進める醍醐味を味わった.その後も,プリオン/アミロイド,非典型的な翻訳動態(新生鎖の生物学)などタンパク質の見方が拡がる研究に従事してきた.これからも未踏のタンパク質世界を開拓していきたい.
小林武彦:東京大学定量生命科学研究所・教授,九州大学大学院医学系研究科博士課程修了(理学博士),米国ロッシュ分子生物学研究所,米国国立衛生研究所,国立基礎生物学研究所・助教授,国立遺伝学研究所・教授などを経て現在に至る.研究テーマはゲノムの維持機構から生命の連続性の謎を解き明かす.伊豆の海と箱根の山をこよなく愛する.著書に「生物はなぜ死ぬのか」(講談社現代新書),「DNAの98%は謎」(講談社ブルーバックス),「寿命はなぜ決まっているのか」(岩波ジュニア新書)などがある.
稲田利文:1987年東京大学理学部生物化学科卒業,’92年東京大学理学系研究科生物化学専攻修了博士(理学),同年名古屋大学理学部分子生物学科助手 ,’98年UC Berkeley visiting scientist(海外学振),2002年名古屋大学理学研究科生命理学専攻准教授,’10年東北大学薬学研究科教授を経て’21年東京大学医科学研究所教授.リボソームを長年研究し,その奥の深さに驚く毎日です.2001年より終止コドンをもたないノンストップmRNAの品質管理の分子機構の解析をしてきました.polyAの翻訳停滞機能,RQCの分子機構の解明,異常翻訳の実体としての衝突リボソームの発見,と翻訳分野が新しいフェーズに変わったことを実感しています.今後もリボソームの隠れた動態制御を明らかにすべく努力したいと思います.