概 論
脂質疾患学アップデート
村上 誠
(東京大学大学院医学系研究科疾患生命工学センター/ 日本医療研究開発機構AMED-CREST)
[略語]
- ABHD:
- α/β hydrolase
- cPLA2:
- cytosolic PLA2
- DHA:
- docosahexaenoic acid
- DRG:
- dorsal root ganglia
- EPA:
- eicosapentaenoic acid
- GPX4:
- glutathione peroxidase 4
- iPLA2:
- Ca2+-independent PLA2
- LDL:
- low-density lipoprotein
- LPA:
- lysophosphatidic acid
- LT:
- leukotriene
- lysoPS:
- lysophosphatidylserine
- NAE:
- N-acylethanolamine
- PAF-AH:
- platelet-activating factor acetylhydrolase
- PAF:
- platelet-activating factor
- PG:
- prostaglandin
- PLA2:
- phospholipase A2
- PLAAT:
- phospholipase A/acyltransferase
- PNPLA:
- patatin-like phospholipase
- sPLA2:
- secreted PLA2
- SPM:
- specialized pro-resolving mediator
はじめに
2010年代から今に至るまで,本邦の脂質研究はまさに黄金期を迎えたといえよう.日本学術振興会新学術領域研究として「生命応答を制御する脂質マシナリー(脂質マシナリー)」(横溝岳彦領域代表,2010〜’14年)と「脂質クオリティが解き明かす生命現象(リポクオリティ)」(有田誠領域代表,’15〜’19年),日本医療研究開発機構AMEDの革新的先端研究開発支援事業CREST/PRIMEとして「疾患における代謝産物の解析および代謝制御に基づく革新的医療技術基盤技術の創出(疾患代謝)」(清水孝雄研究総括,’13〜’20年)と「画期的医薬品等の創出をめざす脂質の生理活性と機能の解明(機能性脂質)」(横山信治研究総括,’15〜’22年)など,脂質にフォーカスを当てた大型競争的研究費が次々と立ち上がり,脂質研究を支えた.わが国の生命科学研究の歴史のなかで,ここまで脂質研究が重要視された時期はこれまでになかったように思う.従来からの脂質研究グループの底上げはもちろんのこと,それまで脂質を扱ったことのなかった多数の研究者も脂質研究に参画し,異分野融合を通じて,まさに脂質を中心に新しい学問が創成されたといえる.本増刊号の前身である「脂質疾患学 なぜ “あぶら” の異常が病気を引き起こすのか?」は,新学術領域研究「脂質マシナリー」が終了したタイミングで2015年に発行された.「脂質マシナリー」は脂質メディエーターに特化した研究領域であり,必然的に2015年の増刊号も脂質メディエーターを中心に構成された.その後8年を経て,一連の上記大型研究の集大成として,続編「治療標的がみえてきた脂質疾患学」を発行することとなった.本増刊号では,「脂質マシナリー」に根ざした脂質メディエーター研究のさらなる展開に加えて,「リポクオリティ」の流れを汲んだ脂質分子の多様性とその意義,脂質の四大機能(細胞膜構成成分,エネルギー源,シグナル分子,体表バリア)を総合的に俯瞰した疾患生物学,オミクス全盛時代に呼応した新しい脂質解析技術の開発,異分野融合により創出された脂質研究の新規概念が幅広くカバーされており,この10年余りの間に脂質研究がいかに飛躍的発展を遂げたかを実感できる内容となっている.
1.本増刊号の構成
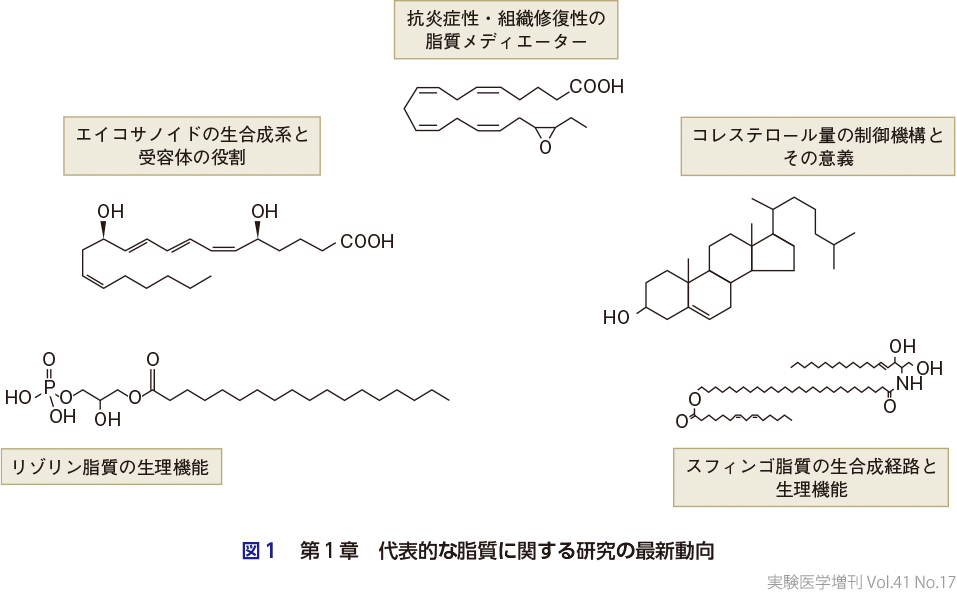
本増刊号は,4つの大項目から構成されている.第1章(図1)は,2015年の増刊号「脂質疾患学 なぜ “あぶら” の異常が病気を引き起こすのか?」を踏襲し,生体における脂質機能とその最新像と称して,代表的な脂質に関する研究の最新動向について,それぞれのエキスパートにまとめていただいた.横溝は,最も代表的な脂質メディエーターであるプロスタグランジン(PG)やロイコトリエン(LT)などのエイコサノイドについて,生合成系と受容体に関するこれまでの知見ならびに臨床展開の現状について簡潔に取りまとめている(第1章-1).可野らはリゾリン脂質としてリゾホスファチジン酸(LPA)とリゾホスファチジルセリン(lysoPS)を取り上げ,LPAの血管形成,線維症,着床における役割,lysoPSによる免疫制御について,受容体の立体構造情報を加味して解説している(第1章-2).有田は,ω3脂肪酸カスケードにより生合成される抗炎症性・組織修復性の脂質メディエーター(SPM, specialized pro-resolving mediator)について,皮膚炎や脂肪肝炎などの制御を例に,最近の研究成果を紹介している(第1章-3).植田は,細胞膜がコレステロールの濃度を厳密に感知し調節するしくみについて,この分野の最新トピックスの1つであるAsterを絡めて解説している(第1章-4).木原はスフィンゴ脂質の多様性と生合成経路について解説しており,なかでも皮膚バリアに必須の脂質であるアシルセラミドの代謝経路の全体像の解明は特筆に値する(第1章-5).
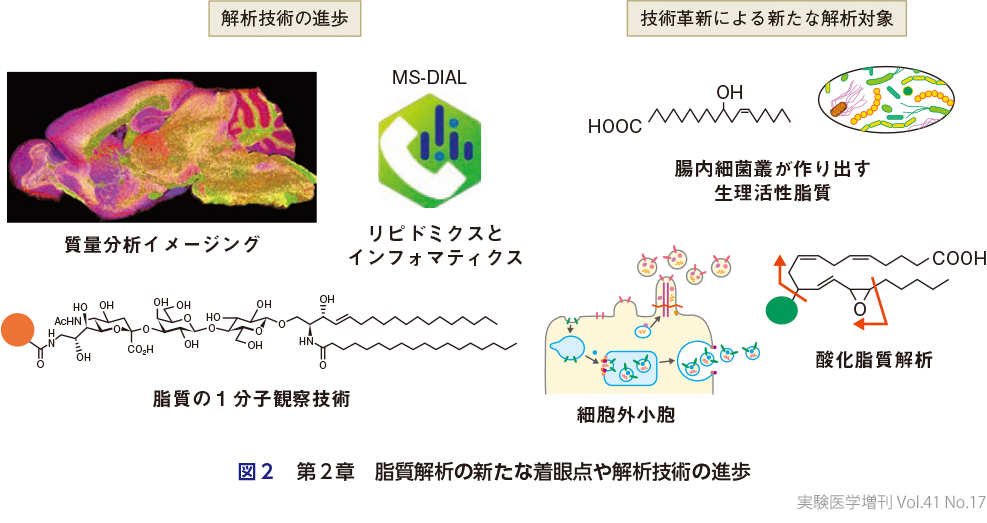
第2章(図2)では,脂質解析の新たな着眼点や解析技術の進歩について解説している.いまや脂質のオミクス解析(リピドミクス)は幅広い研究分野に浸透し,高感度化・スピード化が実現し,トップジャーナルの論文ではリピドミクスの解析データを当たり前のように見かけるようになった.分析する脂質の種類を定めた従来型のターゲット型リピドミクスに加えて,未同定の脂質までも包含したノンターゲット型リピドミクスの技術が開発され,理論上10万種を超えるとされる脂質の網羅的定量分析が可能となり,膨大な情報を統合処理するインフォマティクス技術(MS-DIAL)も開発された(津川・有田,第2章-3).組織切片上で個々の脂質分子の局在を可視化する質量分析イメージングの技術開発も進んでおり,高感度化・高解像度化を通じて,組織中に微量にしか存在しないリゾリン脂質の局在を可視化できるレベルにまで達している(可野ら,第2章-4).このような技術革新の結果として,これまでに解析対象になっていなかった脂質分析も容易となり,本増刊号ではその例として,細胞外小胞の形成過程における脂質膜の変形切断機構(西村・末次,第2章-1),腸内細菌叢がつくり出すユニークな生理活性脂質(渡辺ら,第2章-2),高度不飽和脂肪酸がラジカル酸化されて生じる多様な代謝物(山田,第2章-6)を取り上げている.しかしながら,1細胞レベルやオルガネラレベルでの脂質の質量分析イメージングは現段階では達成されておらず,ライブイメージングも困難である.それを補う技術として,脂質プローブと超解像蛍光顕微鏡を組合わせた脂質の1分子観察技術が開発され,いくつかの方法論と実施例が紹介されている(田口・鈴木,第2章-5).
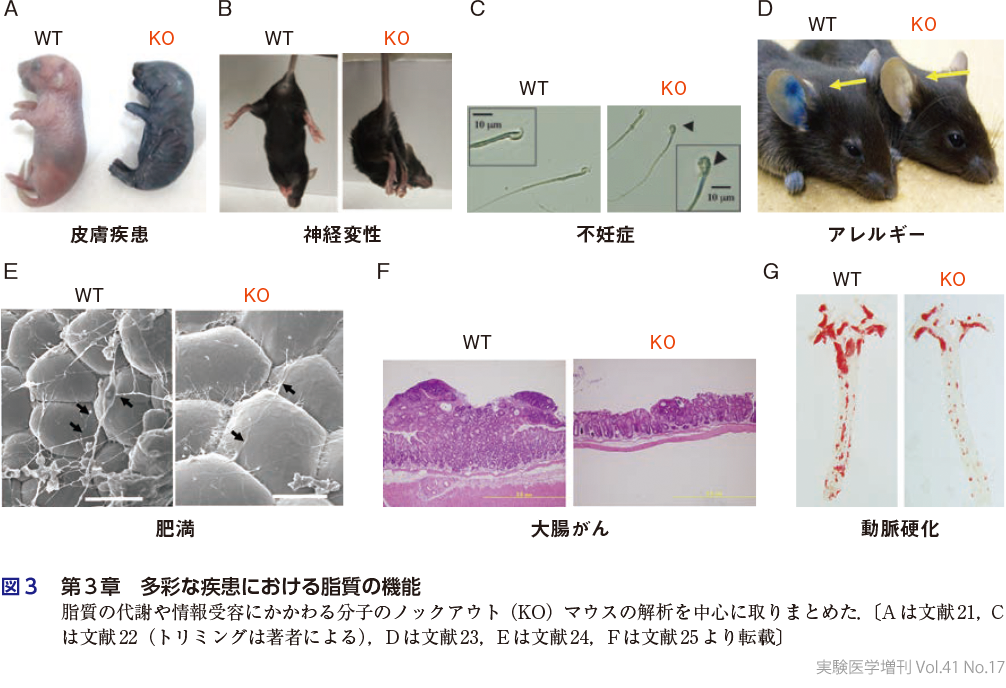
第3章(図3)は「脂質疾患学」の核となる章でもあり,多彩な疾患における脂質の機能について扱っている.がんに関しては,PGE2受容体を標的としたがんの免疫チェックポイント療法の現状と将来展望(土屋・杉本,第3章-1),B細胞リンパ腫形成における細胞外小胞の脂質の役割(幸谷・村上,第3章-2)について取り上げる.免疫疾患については,活性化細胞やアポトーシス細胞におけるリン脂質二重膜のスクランブリングの分子機構(蚊谷ら,第3章-3),T細胞のイムノメタボリズム(遠藤,第3章-4),C型レクチン受容体による糖脂質リガンドの認識とシグナル伝達機構(渡邉・山﨑,第3章-5),喘息におけるシステイニルLTの役割とその創薬展開(永井 潤,第3章-6)などの話題を提供する.神経疾患に関しては,神経ストレスに伴う神経免疫連関における脳内エイコサノイドやSPMの役割(永井裕崇・古屋敷,第3章-7),脳虚血後の修復にかかわる脂質メディエーターの動員機構と作用機序(七田,第3章-8),神経性疼痛の調節因子としての血小板活性化因子(PAF)の新機能(進藤・山本,第3章-9),高度不飽和脂肪酸の欠乏モデルから明らかとなった双極性障害との関連(李・横溝,第3章-10)を取り上げる.脂質異常が肥満・代謝性疾患と関連することは容易に予想がつくが,これと関連する話題として,脂肪酸伸長酵素ELOVL6による脂肪酸長の調節と2型糖尿病・脂肪肝の関連(松坂・島野,第3章-11),ホスホリパーゼA2(PLA2)ファミリーによるリン脂質・中性脂質の新陳代謝と肝疾患の関係(平林・村上,第3章-12),動脈硬化におけるLDL修飾の重要性に関する最新知見(宮崎拓郎・宮崎 章,第3章-13),肥満・脂肪肝や急性腎障害における細胞死に伴う脂質代謝の変容とその意義(菅波ら,第3章-14),PGE2シグナルの遮断による脂肪組織の線維化抑制と健康肥満(稲住・杉本,第3章-15)について解説する.皮膚も脂質の変容の影響を受けやすい臓器であり,前出のセラミドに加えて(木原,第1章-5),イノシトールリン脂質の代謝異常と乾癬,アトピー性皮膚炎の関連(金丸・中村,第3章-16),各種PLA2による脂質メディエーター,バリア脂質,および腸内細菌叢の調節を介した皮膚の変容(武富・村上,第3章-17),各種皮膚疾患におけるエイコサノイドの役割(米倉ら,第3章-18)の多角的視点から,皮膚と脂質の関係について解説する.最後に,雌性不妊(藍川・廣田,第3章-19)と雄性不妊(飯塚・進藤,第3章-20)の観点から,エイコサノイドやドコサヘキサエン酸(DHA)の生殖における意義について解説する.
第4章(図4)では,臨床展開に向けた脂質研究の現状と問題点を探る.蔵野は,疾患バイオマーカーとしてのリゾリン脂質のポテンシャルについて,東京大学病院におけるいくつかの実例(COVID-19患者を含む)をあげて解説している(第4章-1).小田は,さまざまな臨床検体からリピドミクスにより疾患バイオマーカーを同定する試みについて,技術革新と実施例を紹介している(第4章-2).大山らのグループは,浜松医科大学においてわが国最大規模の国際マスイメージングセンターを稼働させており,そこで実施されたヒト検体の空間リピドミクスの事例を紹介している(第4章-3).
2.PLA2研究の今昔物語
さて,ここからは筆者の専門であるPLA2研究の最新動向について,この場を借りて簡単に紹介したい.PLA2関連のトピックスについては本増刊号の各論でもいくつか取り上げられているので(第3章-2, 4, 8, 12, 13, 17),ここでは各論で取り上げられていない話題に触れつつ,これまでの研究の歴史も踏まえて筆者の思うところをざっくばらんに述べてみたい.
1)cPLA2はアラキドン酸の絶対王者か?
PLA2と聞けば,研究者がまず思い浮かべるのはアラキドン酸代謝であり,生化学の教科書にもそのように記載されている.歴史的な流れから,PLA2ファミリーは分泌性PLA2(sPLA2),細胞質型PLA2(cPLA2),Ca2+非依存性PLA2(iPLA2)の三大ファミリー,およびその他のPLA2に大別される(図5).2000年代初期ごろまでは,どのPLA2がアラキドン酸代謝,すなわちエイコサノイドの産生にかかわるかがPLA2研究の中心課題であり,いまでも「PLA2=アラキドン酸代謝」と考える研究者は多い.いうまでもなく,この観点から見たfinal winnerはcPLA2αであり,この酵素がアラキドン酸代謝の中心的な存在であることは疑いの余地はない1).しかしながら,cPLA2αだけでアラキドン酸代謝のすべてを説明できるかというと,そうでもない.PLA2分子群の欠損マウスを網羅的に扱っていると,cPLA2α欠損マウスで,アラキドン酸代謝物であるPGの産生がほとんど低下しないケースや,別のPLA2の欠損マウスでPG産生が低下するケースをたびたび経験する.脳や脂肪組織などいくつかの組織では,リパーゼにより中性脂質から遊離されるアラキドン酸がPG産生と連関している(第3章-1).cPLA2αがその機能をフルに発揮するためには細胞質Ca2+濃度の上昇とリン酸化による翻訳後修飾が必須であり,発現さえしていればアラキドン酸代謝が起こるわけではない.トランスクリプトーム解析でcPLA2α(遺伝子名Pla2g4a)の発現変動を捉えたとしても,それがcPLA2αの活性化と直結しているかはわからない.また,cPLA2αはアラキドン酸に対する特異性が高いため,DHAに由来するSPMの産生をcPLA2αで説明している論文を見かけたら,信用しない方がよい.少し脱線するが,2022年に,EPAやDHAに由来するSPMが生体内において生物学的に意味のある量として存在するかについて,肯定派と否定派にわかれて大激論が交わされた2).個人的には,SPM研究のこれまでの世界的な進展を見れば,「SPMを検出できない」という否定派の意見は的外れに思われるが,PG,LTと比べて産生量がかなり少ないことは確かであり,今後の議論の行方を見守りたい.話をcPLA2に戻すと,cPLA2α以外のアイソフォームの機能は長年不明であったが,最近,cPLA2εがエイコサノイドとは別の脂質メディエーターであるNAE(N-acylethanolamine)の産生にかかわることが明らかとなった(第3章-17).すなわち,cPLA2ファミリー全体を安直にアラキドン酸代謝と結びつける発想は必ずしも正しくなく,アラキドン酸代謝との関連に言及する際には必ずcPLA2 “α” と表記していただきたい.
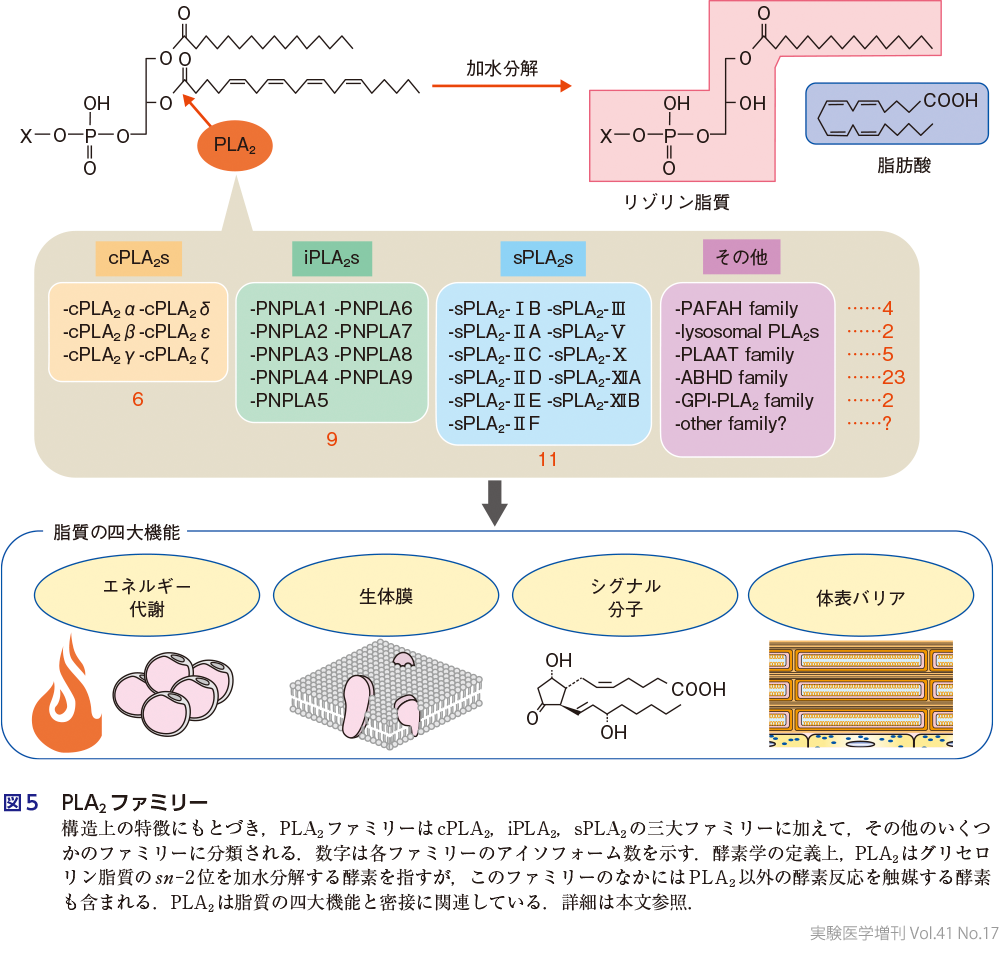
2)sPLA2の機能
sPLA2のバイオロジーはこの十数年の間に飛躍的に発展し,全11種類のアイソフォームの生体内機能が解明された.各sPLA2アイソフォームの機能の詳細については割愛するが,興味のある方は最新の総説を参照されたい3).現在のコンセンサスとしては,sPLA2の標的基質は細胞外に存在するリン脂質であり,細胞外小胞,リポタンパク質,サーファクタントなどの内在性リン脂質のほか,外来成分である食事リン脂質や微生物膜などが含まれる.一部のsPLA2は微生物膜に対する活性がきわめて強く,感染防御因子として作用する一方で,腸内細菌叢の制御を介して二次的に全身の免疫や代謝にも影響をおよぼす.このようなsPLA2の機能は総じて細胞外で発揮されていることから,脂質メディエーターの産生と結びつくケースを除き,脂質の四大機能とは分けて考えるべきかもしれない.われわれの検討によれば,sPLA2が生体内でつくり出す主要な脂質メディエーターはPG,LTよりもむしろリゾリン脂質であり,多くの場合,細胞外小胞が足場となっているようである(第3章-2).一方,sPLA2が細胞膜のリン脂質を基質とするかについては議論の余地がある.強力な活性をもつ蛇毒sPLA2は容易に細胞溶解を引き起こすが,一般に正常細胞の膜は内因性sPLA2に対する感受性が低く,生理的濃度のsPLA2による細胞膜リン脂質の分解は虚血や潰瘍などで細胞がダメージを受けたときにしかみられない(第3章-8).しかしながら最近,敗血症においてsPLA2-Ⅴが赤血球の膜を分解して細胞溶解を引き起こすことが報告され,これは内因性sPLA2が蛇毒sPLA2様の溶血活性を発揮することを示したはじめての事例である4).また,重症COVID-19患者の予後不良とsPLA2-ⅡAの血中濃度の間に強い相関があることが報告されたが5),単なるバイオマーカーなのか,それとも本酵素が “エイコサノイドストーム” に寄与しているかは不明である.
3)iPLA2の機能
iPLA2は別名PNPLA(patatin-like phospholipase)ともよばれ,真核生物に広く種を超えて分布していることから,その機能の本質は「生命応答の根幹をなす脂質代謝」にあると考えられる.実際,iPLA2ファミリーはPLA2活性以外にもリパーゼ,トランスアシラーゼ,リゾホスホリパーゼ活性などを示し,この十数年の間に細胞膜の新陳代謝,エネルギー代謝の調節,脂質メディエーターの産生,バリア脂質の生成などの脂質の四大機能に多様にかかわることが示されてきた1).iPLA2ファミリーの欠損や変異はヒト遺伝性疾患との関連も深く,cPLA2やsPLA2の欠損と比べてはるかに重篤な症状を呈する.ここでは,リン脂質分解型酵素の代表としてPNPLA9(別名PLA2G6またはiPLA2β),中性脂質分解型酵素の代表としてPNPLA2(別名ATGL,adipose triglyceride lipase)の最新の話題を紹介したい.
PNPLA9の変異は乳幼児軸索ジストロフィー,パーキンソン病などの神経変性疾患を引き起こすことが知られており,そのメカニズムとして,脳内DHA含有リン脂質の減少,ドーパミン神経のCa2+シグナル異常,脳内リン脂質の脂肪酸鎖長の変化などが提唱されてきた.最近これに加えて,PNPLA9が過酸化リン脂質の分解除去を介してフェロトーシスを抑えることが報告された6).PNPLA9変異に起因するパーキンソン病では過酸化リン脂質が脳内に蓄積し,ドーパミン神経がフェロトーシスを起こして死滅する.この経路は,過酸化リン脂質消去の中心酵素であるGPX4とは異なる新たなフェロトーシス制御機構を提示するものである.
PNPLA2はトリグリセリドの分解に必須のリパーゼであり,この十数年の間に,その複雑かつ巧妙な調節機構が解き明かされた(第3章-12参照).脂肪滴形成過程において,新規合成されたトリグリセリドは脂肪滴に蓄えられるが,同時に脂肪分解も活発に起こっている.このエネルギー的に高コストに思われる脂肪合成(lipogenesis)と脂肪分解(lipolysis)の無益回路(futile cycle)は,一説には体温維持のための熱産生に利用されるといわれているが,これに加えて新たな説が提唱された.細胞膜のリン脂質と同様に,脂肪滴のトリグリセリドも分子種レベルで活発に脂肪酸リモデリングを生じており,脂肪分解を担うPNPLA2と脂肪合成を担うDGAT1/2の連携により,トリグリセリドの3本の脂肪酸鎖が数時間のうちに完全に入れ替わる.つまり,総量としてのトリグリセリドは変わらないように見えても,分子種レベルとしてのトリグリセリドは急速に変化しており,細胞内の代謝要求性に応じて脂肪滴から動員される脂肪酸の種類を変化させているのである7).この意味で,PNPLA2は単にβ酸化の基質としての脂肪酸を供給するだけでなく,トリグリセリドの質(リポクオリティ)の制御にかかわる鍵分子ともいえる.2本の脂肪酸がエステル結合で結ばれた分岐脂肪酸(FAHFA,fatty acid ester of hydroxy fatty acid)は,脂肪酸受容体GPR120を介して代謝改善・抗炎症作用を示す新しいタイプの生理活性脂質であるが,その生合成機構は不明であった.ごく最近,PNPLA2がトランスアシラーゼ活性を介して分岐脂肪酸の生合成にかかわることが明らかとなった8).さらに網膜において,PNPLA2は網膜上皮細胞の脂肪滴に蓄えられたレチニルエステルを加水分解して11-cis-レチナールを動員し,視機能の調節にかかわることが報告された9).この発見に先立って,PNPLA2が網膜のリン脂質を分解してDHAを遊離すると報告されていたが,PNPLA2はPLA2活性をほとんど示さないので,これはPNPLA2の別名であるiPLA2ζの名称に惑わされた誤認であると筆者は考えている.
4)その他のPLA2
その他のPLA2については各論では全く触れられていないので,いくつかホットな話題を紹介したい.PAF-AH(PAF acetylhydrolase)は,その名の通りPAFを分解する酵素として同定されたが,生理的には酸化リン脂質を選択的に分解するPLA2と考えられている.このうち,血漿型PAF-AH(一般名PLA2G7)は低密度リポタンパク質(LDL)に結合した形で存在することからLp-PLA2ともよばれ,動脈硬化との関連が注目されてきた.動物実験において本酵素の特異的阻害剤Darapladibの抗動脈硬化薬としての有効性が示されたが,臨床試験で頓挫した10).一方,PLA2G7欠損が加齢に伴う炎症と代謝異常を改善することが見出され,本酵素と老化の関連が改めて見直されている11).また,PLA2G7はDRGニューロンに最も高発現しているPLA2であり,欠損マウスではω6脂肪酸の過剰摂取による神経性疼痛の増悪が抑えられる12).この論文では,PLA2G7により遊離されたω6脂肪酸が病態増悪に寄与すると結論づけているが,本酵素の基本的性質を加味すると,ω6脂肪酸そのものではなく,その酸化代謝物をLDLリン脂質から遊離している可能性が高い.PLA2G7と近縁の細胞質酵素であるPAF-AH2は,細胞膜リン脂質からω3脂肪酸代謝物(ω3エポキシド)を直接遊離してマスト細胞の活性化を増強するとともに13),肺高血圧症の悪化の要因となる肺血管周囲の線維化を抑える14).
リソソーム型PLA2として知られるPLA2G15(別名LPLA2)は,リソソーム内に取り込まれたリン脂質をPLA1/2活性により消化すると考えられてきた.最近,PLA2G15によりリソソーム内で生じたリゾリン脂質が,リソソーム膜に存在するリゾリン脂質輸送体Spns1により細胞質に汲み出され,小胞体において膜リン脂質のリモデリングに利用されることが判明した15).この論文によれば,リソソームに由来するリゾリン脂質のプールは予想以上に大きく,またリソソームは細胞外から取り込んだ脂質粒子だけでなくオートファジーを通じて細胞内オルガネラ膜の分解にも関与するため,今後の膜リン脂質リモデリング研究においては,リソソームにおいてPLA2G15により産生されたリゾリン脂質の寄与も考慮に入れる必要があろう.
さらに最近,オートファジーに依存しないオルガネラ膜の大規模分解機構が存在することが判明した.PLA2G16(別名PLAAT3)は,もともと脂肪細胞特異的に発現している新しいタイプのPLA2として同定された.この酵素の欠損マウスは脂肪組織が萎縮し,肝臓に異所性脂肪が蓄積してlipodystrophyの表現型を呈する16).そのメカニズムとして,PLA2G16により遊離されたアラキドン酸がPGE2に代謝されてEP3経路を活性化するためと半ば強引に説明されたが,EP3受容体の欠損マウスはlipodystrophyの表現型を示さないので,別の可能性を考えるべきである.時を経て,PLA2G16が細胞内オルガネラ膜の大規模分解を通じて水晶体レンズの透明化にかかわることが明らかとなった17).すなわち,PLA2G16は脂質二重膜の何らかの性状変化を認識してオルガネラ膜に突き刺さり,PLA1/2活性によりオルガネラが粉々になるまでリン脂質を分解する.PLA2G16によるオルガネラ膜の完全分解が水晶体以外の組織でも起こるかどうかについては未解明であるが,この発見はPLA2ファミリーの新しい作動原理の解明につながるものである.さらに最近,PLA2G16が属するPLAATファミリーの別の一員であるPLAAT1が,おそらくPLA1/2として脂肪肝の増悪にかかわることが明らかとなった18).
ABHDファミリーは新たに認識されたPLA2ファミリーの一群であり,このなかにはミリスチン酸含有リン脂質を特異的に分解するABHD1とABHD3,3本の脂肪酸をもつN-アシルリン脂質を分解するABHD4,酵素活性をもたないがPNPLA2の補助因子としての機能をもつABHD5,1本の脂肪酸をもつリゾリン脂質やモノグリセリドを分解するABHD6,lysoPSを選択的に分解するABHD12,PSを分解するABHD16A,脱パルミトイル化酵素として働くABHD17など,興味深い特徴を示す酵素が含まれている19).ABHD12は多発性神経炎,難聴,運動失調,網膜色素変性,白内障を伴うPHARC症候群,ABHD16Aは遺伝性痙性椎麻痺の原因遺伝子である20).このファミリーはまだ文献が少ないため,提唱されているメカニズムにはまだ曖昧な点が残されており,今後の研究による新しい展開に期待したい.
最後に,論文未発表のため詳述は避けるが,PLA2ファミリーにはまだ残された隠し球が存在するらしい.PLA2研究は実に奥が深い.これだけ多くのPLA2が存在する理由はなぜだろうか?この疑問に対する明確な答えはないが,それだけ多様で複雑な脂質代謝経路が生体内に存在し,それに適材適所で対応するために多数のPLA2が分子進化してきたことは間違いない.
おわりに
多少散漫な内容となったが,本増刊号の流れについて簡単に概説した.詳しくは,迫力満点の各論をぜひお読みいただきたい.単純に脂質研究といっても膨大な広がりがあり,さらに異分野との融合を通じて,脂質生物学はますます複雑化・多様化している.これに対応するための技術革新が飛躍的に進んだことは間違いないが,まだ途上であることもまた事実である.それぞれの各論において,これまでに何が明らかとなって,今後の宿題として何が残されているのかを読みとっていただければ幸いである.2015年発行の前号「脂質疾患学 なぜ “あぶら” の異常が病気を引き起こすのか?」をおもちであれば,それと比較していただくとよい.前号から本号までの8年間に,いかに脂質研究が飛躍的に発展したかを理解していただけるはずである.
文献
- Murakami M:Pharmacol Ther, 244:108382, doi:10.1016/j.pharmthera.2023.108382(2023)
- Sinha G:Science, 376:565-566, doi:10.1126/science.abq8310(2022)
- Murakami M, et al:Immunol Rev, doi:10.1111/imr.13205(2023)
- Takahama M, et al:bioRxiv, doi:10.1101/2023.01.30.526342(2023)
- Snider JM, et al:J Clin Invest, 131, doi:10.1172/JCI149236(2021)
- Sun WY, et al:Nat Chem Biol, 17:465-476, doi:10.1038/s41589-020-00734-x(2021)
- Wunderling K, et al:Nat Metab, 5:699-709, doi:10.1038/s42255-023-00769-z(2023)
- Patel R, et al:Nature, 606:968-975, doi:10.1038/s41586-022-04787-x(2022)
- Hara M, et al:Cell Rep, 42:112091, doi:10.1016/j.celrep.2023.112091(2023)
- Wilensky RL, et al:Nat Med, 14:1059-1066, doi:10.1038/nm.1870(2008)
- Spadaro O, et al:Science, 375:671-677, doi:10.1126/science.abg7292(2022)
- Boyd JT, et al:Nat Metab, 3:762-773, doi:10.1038/s42255-021-00410-x(2021)
- Shimanaka Y, et al:Nat Med, 23:1287-1297, doi:10.1038/nm.4417(2017)
- Moriyama H, et al:Nat Commun, 13:3013, doi:10.1038/s41467-022-30621-z(2022)
- He M, et al:Proc Natl Acad Sci U S A, 119:e2210353119, doi:10.1073/pnas.2210353119(2022)
- Jaworski K, et al:Nat Med, 15:159-168, doi:10.1038/nm.1904(2009)
- Morishita H, et al:Nature, 592:634-638, doi:10.1038/s41586-021-03439-w(2021)
- Rahman SMK, et al:FASEB J, 37:e23032, doi:10.1096/fj.202201033R(2023)
- Bononi G, et al:J Med Chem, 64:9759-9785, doi:10.1021/acs.jmedchem.1c00624(2021)
- Lemire G, et al:Am J Hum Genet, 108:2017-2023, doi:10.1016/j.ajhg.2021.09.005(2021)
著者プロフィール
村上 誠:長野県松本市生まれ.1986年東京大学薬学部卒業,’88年東京大学大学院薬学系研究科修士課程修了,’91年東京大学大学院薬学系研究科博士課程修了.’91〜’92年日本学術振興会特別研究員.’93〜’94年米国ハーバード大学留学.’95〜’97年昭和大学薬学部講師,’98〜2004年同助教授.’05〜’10年東京都臨床医学総合研究所副参事研究員,’11〜’16年東京都医学総合研究所参事研究員.’17年より東京大学大学院医学系研究科教授(現職).1999年日本薬学会奨励賞,2000年日本炎症再生医学会奨励賞,’14年テルモ財団賞,’23年文部科学大臣賞.研究テーマ:PLA2分子群の総合的機能解析.趣味:愛犬と戯れること.