
概論
疾患研究に新たな切り口をもたらすタンパク質発現異常の新メカニズム
田口英樹(東京科学大学総合研究院細胞制御工学研究センター),松本有樹修(名古屋大学大学院理学研究科)
はじめに
生命のセントラルドグマ周辺の研究が進めば進むほど,DNAからRNAへの転写,RNAからタンパク質への翻訳の各段階は精巧に調節されていることがわかってきた.タンパク質が合成されてくる直接の過程は翻訳であるが,タンパク質の発現を考えるにあたって,転写を抜きにして議論するわけにはいかない.そこで本増刊号では,さまざまなレベルでのタンパク質の発現異常を,転写・翻訳という2つの過程とRNA・タンパク質という2つの物質に切り分けて紹介することとした.
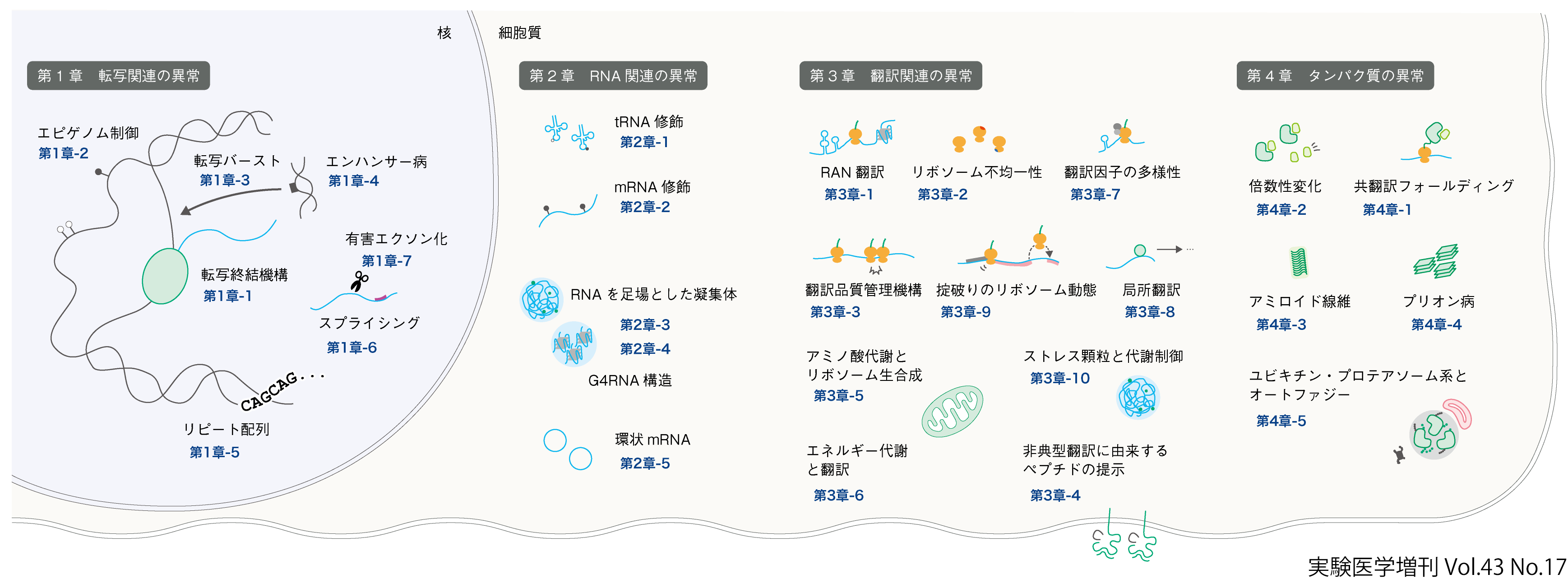
第1章 転写関連の異常編集担当:松本有樹修
最初の段階である転写は遺伝子発現量を決定する要である.この転写制御が乱れると,産生されるタンパク質の量やタイミングが異常となり,細胞機能の破綻や疾患発症につながる.近年,網羅的な転写産物解析プロジェクト(FANTOMやENCODEなど),エピゲノム解析手法,単一細胞レベルのライブイメージング,ロングリードシークエンシング技術の進展により,転写制御の多様なしくみとその破綻メカニズムが次々に明らかになってきた.
中山・野島は転写終結機構の多様性とその破綻が細胞機能に及ぼす影響を論じる.mRNAの適切な発現には転写伸長と終結の均衡が不可欠であり,未成熟終結やウイルスによる制御破綻はゲノム不安定化や病態を招く.タンパク質コード遺伝子と非コードRNAにおける終結様式の違い,未成熟転写終結を利用した新たな細胞機能調節機構について最新の知見が紹介される(第1章-1).
堀居・井上はDNAメチル化やヒストン修飾といったエピゲノム制御の異常に注目し,その破綻が遺伝子発現や発生異常をもたらすしくみを解説する.典型的な刷り込み遺伝子の制御異常による疾患の例に加え,H3K27me3による非典型的刷り込みの破綻が胎盤異常や致死を招くこと,さらにエピゲノム編集による疾患モデル作製の試みが紹介される(第1章-2).
深谷は個体発生における転写バーストの制御機構を論じる.ライブイメージング解析により,エンハンサーが転写バースト頻度を調整する主要因であることが示され,さらにコアプロモーター配列,複数エンハンサーの拮抗作用,eRNA転写や転写因子クラスター形成が多階層的に関与することが明らかとなった.これら制御機構の破綻は体節形成異常など発生異常につながる(第1章-3).
村上・孫はエンハンサー領域の変異が疾患発症に与える影響を取り上げる.自己免疫疾患に関連するSNPの多くがエンハンサーに存在し,遺伝子発現量の異常を通じて病態を引き起こすと考えられる.本稿ではエンハンサー病の概念と,その同定に有効なNET-CAGEやReapTECなどの技術開発,それらを用いた免疫疾患解析の成果が紹介される(第1章-4).
宮武・松本直通はリピート配列の異常伸長が神経筋疾患をはじめとする多彩な病態に関与することを論じている.伸長リピートは多様なレベルで転写制御に障害を与え,エピゲノム変化,RNA foci形成,スプライス異常やRAN翻訳による毒性などをもたらす.近年はロングリードシークエンスにより新規疾患の同定や治療標的探索が加速している(第1章-5).
長谷川らはRNAスプライシングの分子機構とがんにおける異常を取り上げる.スプライシングは多様なアイソフォーム生成を担う重要な過程であり,その破綻は腫瘍発生や薬剤耐性に深く関与する.SF3B1やZRSR2などスプライシング因子変異が造血腫瘍で機能的意義をもつこと,異常産物がNMDを介して分解されるしくみ,さらには治療標的としての可能性が示されている(第1章-6).
芳本はイントロン由来の隠れエクソンが誤って取り込まれる有害スプライシングと,その抑制機構に焦点を当てる.霊長類ではhnRNP CがAlu配列のエクソン化を抑制するのに対し,齧歯類では4.5SH RNAがB1配列の有害エクソン化を防ぐ内在性因子として機能する.4.5SH RNAは標的配列認識と抑制因子リクルートの二重モジュール構造をもち,スプライシング制御に必須であることが示された(第1章-7).
本章では転写異常の観点からタンパク質発現異常疾患の原因解明に迫る最新知見を概観した.転写制御機構の理解深化と技術革新の相乗効果により,疾患における遺伝子発現異常の全貌解明がさらに進み,疾患のさらなる理解につながることが期待される.
第2章 RNA関連の異常編集担当:松本有樹修
RNAはその動的な修飾,立体構造形成,細胞内局在,分解制御などを通してタンパク質発現の微細な調節を担っている.ゆえにRNAに起因する異常はタンパク質発現の破綻を招き,結果として疾患の原因となりうる.実際,tRNAやmRNAの化学修飾異常と疾患との関連,RNAの異常な高次構造形成や凝集体形成,RNAの局在異常や分解経路の破綻など,RNA関連の多彩な異常が疾患と結びつくことが近年明らかになりつつある.
鈴木はtRNA修飾異常と疾患のかかわりを論じている.tRNAは多数の化学修飾を受け,その欠失は翻訳精度や効率を損なう.特にミトコンドリアtRNAの修飾異常はMELASやMERRFなどの病態を引き起こすことが示され,臨床応用に至った例もある.また細胞質tRNAの修飾異常は神経疾患や糖尿病,がんと関連し,tRNA由来断片の機能にも影響を与える(第2章-1).
王はmRNA修飾の一種であるm6Aメチル化が神経細胞における遺伝子発現の時空間的制御に果たす役割を論じている.m6AはmRNAの局在や安定性,翻訳効率に影響し,シナプス可塑性や神経回路形成を支える局所翻訳を調整する.m6Aが神経発達障害や精神疾患にかかわることも示唆されてきており,読み取りタンパク質や修飾酵素を介した制御機構の重要性が明らかになりつつある(第2章-2).
山崎・廣瀬はRNAが足場となって形成される生体分子凝縮体に注目し,その破綻が疾患につながるしくみを解説する.arcRNAによるパラスペックルやストレス顆粒の形成は遺伝子発現制御やストレス応答に重要であり,リピートRNAやXISTなどの異常は神経変性疾患,筋疾患,自己免疫疾患などを引き起こす.疾患関連RNAによる凝縮体形成と,それを標的とした創薬戦略の展望が紹介される(第2章-3).
矢吹・塩田はプリオン性タンパク質の凝集機構におけるRNAグアニン四重鎖の役割を明らかにしている.αシヌクレインやタウは液-液相分離を介してゾル-ゲル相転移を起こし,神経変性に至る.G4RNAが凝集の足場として作用し,シナプス関連mRNAとともに隔離されることで機能障害を引き起こすことが示される(第2章-4).
福地らは環状mRNAが高い安定性をもつ一方で翻訳効率がきわめて低いという課題に対し,Internal Cap-Initiated Translation(ICIT)機構を開発した.枝分かれ型と貼り付け型の二方式によりキャップ構造を導入することで,高い翻訳活性と修飾核酸導入の両立を実現し,マウスにおける持続的発現も確認された(第2章-5).
本章を通じて,RNAの修飾・構造・凝集などの異常がタンパク質発現異常疾患の発症機構に深く関与する現状が浮き彫りとなり,これらの分子基盤の解明が新たな疾患治療法の創出につながる可能性が示唆されている.
第3章 翻訳関連の異常編集担当:田口英樹
DNAにコードされた遺伝情報がリボソームでRNAを介してアミノ酸配列に変換される翻訳では,mRNAの開始コドン(AUG)から翻訳が開始して,コドンに従ってアミノ酸が連結して終止コドンで終了する,というのが基本である.しかし,このような標準的なしくみに従わない翻訳が多々見つかってきており,種々の疾患に関与することがわかってきた.このような新常識が明らかになった背景にはリボソームプロファイリング(Ribo-Seq)などの技術革新,液-液相分離など細胞内のタンパク質の見方のアップデートなどもかかわる.個別に見てみよう.
まず,従来は翻訳されないと思われていたmRNAからの翻訳がさまざまなところで起こっていることがわかってきた.藤野・永井には神経変性疾患における非AUG翻訳について解説してもらった(第3章-1).古くから遺伝性の神経変性疾患において塩基リピートの異常な伸長が知られており,ノンコーディングリピート病と総称されていた.これまでは主にリピートRNAそのものの毒性に焦点が置かれていたが,2011年にAUGをもたない領域から翻訳が起こるRepeat-Associated Non-AUG翻訳(RAN翻訳)が発見された.RAN翻訳では想定される読み枠すべてから翻訳が起こることも興味深い.金関には,T細胞免疫系において重要なMHCクラスⅠに提示される膨大な種類のペプチドの全貌(免疫ペプチドーム)を論じてもらった(第3章-4).免疫ペプチドームは既知のORFに由来する通常のプロテオームに由来するとは限らず,非典型的な翻訳に由来するペプチド(cryptic peptide)が多数含まれている.cryptic peptideが「がん抗原」のソースになっている可能性もあるらしい.近年では,非典型的な翻訳に由来するタンパク質やペプチドが膨大になるらしいということがRibo-Seqや質量分析によるプロテオミクスで明らかになっており,総じて「ダークプロテイン」,「ダークプロテオーム」という名称が定着しはじめている.
翻訳における主役のリボソームや翻訳因子についても新たな展開がある.リボソームはリボソーマルRNA(rRNA)と80種類にも及ぶリボソームタンパク質からなるが,その組成が「不均一」であることがわかってきた.白石・松本有樹修は,リボソーム不均一性の解説とともに,彼らが発見した心臓に特異的に発現するリボソームとその翻訳特異性なども紹介している(第3章-2).翻訳因子も標準以外の因子の活躍が続々と明らかになり目が離せない状況である.例えば,國富・山中は山中が1997年に発見したeIF4G2(NAT1)というeIF4G1のオルタナティブ因子が,機能解析を進めると予想以上に広範かつ重要な働きをしていることを紹介している(第3章-7).今後の翻訳開始の常識が大きく変わる可能性を秘めている.
翻訳は細胞内で最もエネルギーを使うプロセスであり,哺乳類細胞全体では20〜30%のエネルギーを消費するという見積りがあり,種々の代謝制御と密接にかかわる.大澤はがん細胞を例としたアミノ酸代謝とリボソーム生合成について解説している(第3章-5).細胞内のエネルギー産生はミトコンドリアに担われており,大量にエネルギーを使う翻訳と共役していることがわかってきた.勝村・森田は,ミトコンドリアと翻訳の関係,特にmTORC1シグナル伝達経路などについて紹介している(第3章-6).
翻訳を司るリボソームの動態もかなり多様であることがわかってきた.近年の細胞内タンパク質の恒常性(プロテオスタシス)分野のホットトピックスの一つ,翻訳品質管理機構(Ribosome-associated Quality Control:RQC)のパイオニアである稲田は,翻訳速度の低下などによって起こる衝突リボソームとストレス応答などの関連について解説してくれた(第3章-3).茶谷は,フレームシフトや翻訳一時停止など掟破りのリボソーム動態を詳しく論じるとともに,これらの掟破りが遺伝情報発現の拡張にもつながっている例を紹介している(第3章-9).
また,翻訳が細胞内のどこで行われるのか,さらに,その制御も近年明らかになりつつある.七野はmRNAやリボソームが細胞内の特定領域に局在してタンパク質を合成する局所翻訳と,その神経変性疾患との関連を解説してくれた(第3章-8).また,七野は局所翻訳を解析する重要な手法として本増刊号でたびたび登場するRibo-Seqにも言及してくれた.翻訳される場所という観点から,細胞内での液-液相分離現象を避けるわけにはいかない.武川はストレス顆粒という相分離でできる典型的な非膜オルガネラでは,翻訳抑制のみならず,RNA代謝制御など種々の生命現象が調節されている知見をまとめてくれた(第3章-10).
第4章 タンパク質の異常編集担当:田口英樹
最終章は翻訳以降,実際にはリボソームでmRNA上のコドンが読まれて誕生するところから死に至る(分解される)までの「タンパク質の一生」を扱う.まずは,タンパク質の発現量に関する不均衡について取り上げた.上原には,さまざまな発生異常やがん形成などにかかわる倍数性変化について解説していただいた(第4章-2).倍数性が変化した細胞では遺伝子量の不均衡が,mRNAやタンパク質の量の不均衡につながり,タンパク質毒性ストレスを引き起こす.
タンパク質の立体構造形成,すなわちフォールディングの異常もさまざまな疾患にかかわる.田口は翻訳に共役したフォールディング(共翻訳フォールディング)に関して概説した(第4章-1).新生ポリペプチド鎖は翻訳途上でもフォールディングを開始するが,翻訳一時停止など翻訳動態に依存してフォールディングの状況が変化しうる.究極的には,アミノ酸配列が同一の同義置換で翻訳速度の緩急が影響を受けて,フォールディングの善し悪しが決まるケースがある.これはタンパク質科学の前提である「アミノ酸配列さえ決まればフォールディングは一義に決まる」というAnfinsenのドグマに抵触する.
アミノ酸配列が同一でも異なる立体構造を取るのはアミロイドやプリオンが関与する神経変性疾患でも同様である.佐野・田中には,タウやαシヌクレインを中心としたアミロイド線維の形成・伝播メカニズム,アミロイドの構造多様性について解説してもらった(第4章-3).堀内には,ヒトのクロイツフェルト・ヤコブ病や牛海綿状脳症(いわゆる狂牛病)に代表されるプリオン病の最新知見,特に細胞内増殖や細胞間伝播機構について詳述してもらっている(第4章-4).
細胞内のタンパク質の一生の最終は分解系である.坂巻・小松にはユビキチン・プロテアソーム系とオートファジーによる細胞内タンパク質品質管理機構とその破綻について解説してもらった(第4章-5).

おわりに
これまでに本増刊号のような切り口でタンパク質発現のメカニズム,破綻,疾患との関連を一冊にまとめたような成書はなかったであろう.一つには,これまでの生命科学の大きな成果がセントラルドグマの前半,特に複製や転写制御に寄っていたことにあるのではなかろうか.技術的にも,1980年代には確立したDNA配列決定法が大きな貢献をしたと言える.タンパク質に関しては,2000年代になってようやく質量分析(MS)によるプロテオミクスが実用化され,2010年頃のRibo-Seqで翻訳動態が網羅的に解析できるようになったことが大きく,翻訳レベルでの遺伝情報発現制御の重要さを浮き彫りにしてくれた.とは言え,MS依存のプロテオミクスもRibo-Seqもまだ限界がある.今後,タンパク質関連の画期的な解析技術が産まれることでさらに大きなブレイクスルーがあると思われる.
<著者プロフィール>
松本有樹修:名古屋大学大学院理学研究科 教授.2011年九州大学医学系学府博士課程修了.博士(医学).’11〜’12年同所属学術研究員,’12〜’17年Harvard Medical School博士研究員,’17〜’23年九州大学准教授,’23年から現所属.ダークプロテオーム(非典型的タンパク質)とその発現を制御する翻訳機構の理解を通して疾患の治療などに応用することをめざしている.
田口英樹:東京科学大学総合研究院 教授.1993年東京工業大学総合理工学研究科博士課程修了.博士(理学).’95〜2003年東京工業大学資源化学研究所助手,’03〜’10年東京大学新領域創成科学研究科准教授,’10〜’16年東京工業大学生命理工学研究科教授,’16年から現所属.新しいアイデア,材料,手法を使って細胞内のタンパク質の一生,特にリボソームでの「誕生」やシャペロンによる「自己実現アシスト」について分子機構を解明したい.