
當瀬規嗣
(札幌医科大学医学部細胞生理学講座)
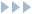
- 腎小体は毛細血管である糸球体と,ボーマン嚢からなる.
 1
1 - 輸入細動脈から流入した血液から糸球体濾過により原尿がつくられる.1, 2
- 糸球体濾過の原動力は血圧であり,阻害要因は膠質浸透圧差とボーマン嚢内圧である.3
- 糸球体の通りやすさは糸球体壁の細胞間間隙によって左右される.4
- 尿細管糸球体フィードバックにより糸球体濾過量(GFR)は調節される.5
 腎小体の構造
腎小体の構造
▶structure of renal corpuscle
腎小体は毛細血管が屈曲しながらたたまれた状態になっている糸球体と,糸球体を包み込むようにしているボーマン嚢からなり立っている(図1).糸球体の壁から原尿が滲みだして,これをボーマン嚢が受け止めて,尿細管へ導くのである.
腎臓において腎小体が存在する領域を皮質とよぶ.皮質の内側は尿細管と血管が豊富にあり,髄質とよばれる.
腹部大動脈から直接分岐した腎動脈は左右の腎臓に流れ込み,腎臓内で枝分かれして,細動脈レベルの太さの輸入細動脈となる.これが糸球体に血液を流し込む.糸球体を通過した血液は輸出細動脈に流れ出て,同じネフロンの尿細管の伴走血管となる.
 糸球体の壁
糸球体の壁
▶wall of glomerulus
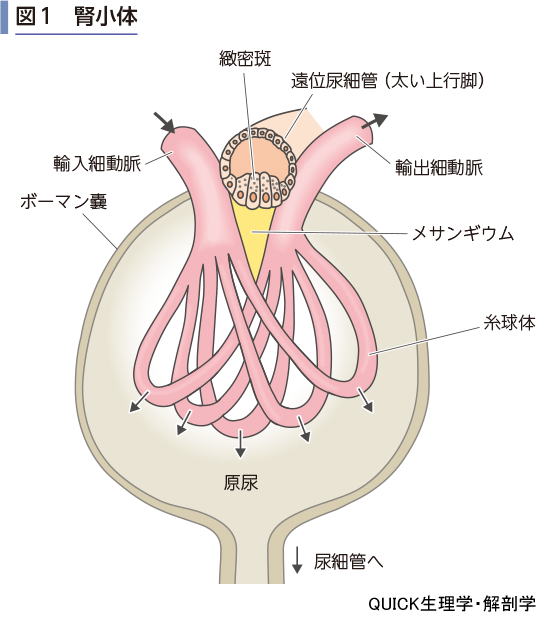
糸球体の壁は他の毛細血管と同じように血管内皮細胞,基底膜で構成されるが,それに加えて血管の外周を上皮細胞(タコ足細胞,ポドサイト)が覆っている(図2).内皮細胞,上皮細胞ともに細胞間接着が弱く,一定程度の間隙がある.したがって,血流があり,血圧が壁にかかるとその力により血漿は糸球体外へしみ出る.血球や,グロブリン,リポタンパク質(キロミクロン,VLDL,LDL,HDLなど)のような高分子タンパク質は,糸球体の間隙を通過できないので,血管内に残る(限外濾過).この過程を糸球体濾過という.こうして原尿がボーマン嚢内にしみ出る.原尿の生成量,すなわち糸球体濾過量(GFR)は両腎の合計で110~135 mL/minにのぼる.
 糸球対濾過の原動力
糸球対濾過の原動力
▶driving force of glomerulus filtration
糸球体濾過の原動力は糸球体内圧すなわち血圧である.圧力の面から考えると糸球体濾過を阻害する要因は2つある.1つは糸球体内外の膠質浸透圧差である(図2).前述したように,高分子タンパク質は濾過されないので,糸球体内側の膠質浸透圧が高いことになる.これが水を糸球体内に引き戻す効果を示す.また,ボーマン嚢内圧が上昇すると,濾過がしにくくなる.ボーマン嚢内圧は,尿の排出経路に障害があると高まる可能性がある.例えば,尿路結石,尿路感染症,腎炎,腎がんなどである.
したがって,全身の血圧が低下すると糸球体濾過量は減少する.特に収縮期血圧が90 mmHgを下回ると,急激に糸球体濾過量は減る.腎機能の維持には血圧の確保が必須なのである.
 糸球体壁の透過性
糸球体壁の透過性
▶permeability in wall of glomerulus
糸球体濾過量を左右する要因のもう1つは,糸球体壁の水の通りやすさで,糸球体濾過係数で示される.ネフローゼ症候群では,糸球体の障害によって糸球体壁の間隙が拡大し,高分子タンパク質も原尿に流出するようになる.この場合,糸球体濾過係数は増加していて,糸球体濾過量が増大し,多尿,タンパク質喪失が起こる.一方,糸球体腎炎では,免疫複合体が糸球体壁に沈着して間隙が詰まってしまう.この場合は糸球体濾過係数が著しく低下し,糸球体濾過量は減少して乏尿となる.
 尿細管糸球体フィードバック
尿細管糸球体フィードバック
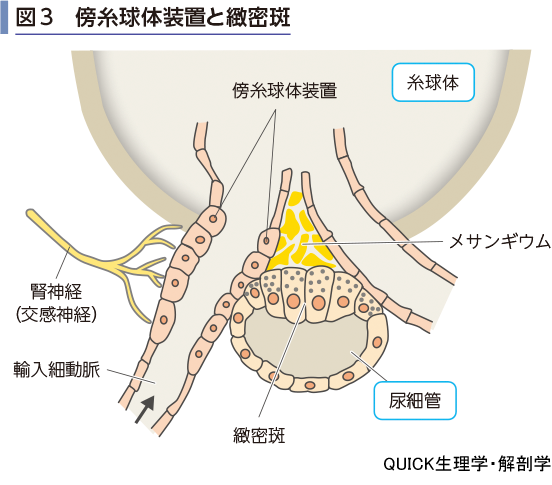
▶tubuloglomerular feedback
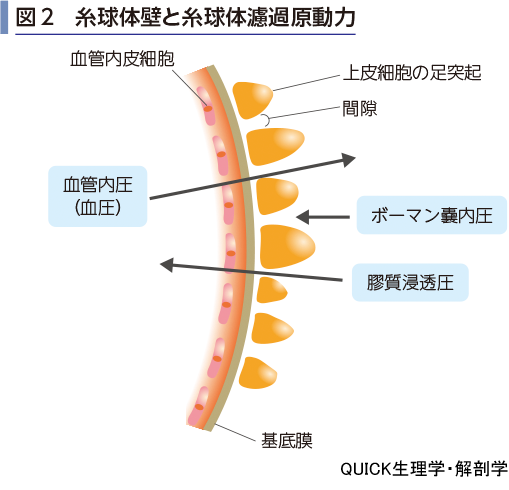
腎小体の輸入細動脈と輸出細動脈の間には,複数の細胞で構成される,重要な領域がある.輸入細動脈の内皮細胞の一部は,背が高い細胞となり,傍糸球体装置とよばれる(図3).ここからはレニンが分泌される(第7章-05-図8参照).傍糸球体装置には,レニン分泌を刺激する腎神経(交感神経性)が伸びてきている.一方,腎小体を発した遠位尿細管が輸入細動脈と輸出細動脈の間を通過するが,その隙間を埋めている細胞をメサンギウムとよぶ.
糸球体濾過を腎臓内で調節するしくみとして,尿細管糸球体フィードバック機構があげられる.遠位尿細管は皮質へとループして起源の腎小体の輸入細動脈と輸出細動脈の間を通り抜けている.細動脈に接する側の遠位尿細管上皮は背の高い細胞となっており,緻密斑とよばれる.この細胞は尿のNaCl濃度を感知している.もし,NaCl濃度が上がると輸入細動脈を刺激して収縮させGFRは低下する.逆にNaCl濃度が低下すると,輸入細動脈が拡張し,糸球体への流入血液量が増えるのでGFRが高くなるのである.
 ネフローゼ症候群
ネフローゼ症候群
高度なタンパク質尿,低タンパク質血症,それにもとづく下肢などの浮腫を主要徴候とする疾患.糸球体濾過のしくみが障害されて,糸球体濾過係数が上がり,高分子であるタンパク質分子(主にアルブミン)が尿へ漏出する.とりわけネフローゼ症候群の1つである微小変化群では糸球体の構築自体は崩れていないにもかかわらず,タンパク質が漏出していることから,細胞間間隙のサイズがタンパク質分子の通過を規定している限外濾過という考えを支持している.

- 糸球体で濾過されないのはどれか.(創作問題)
- aアミノ酸bフルクトースcグルコースdグロブリンeカリウム
- 解答と解説はWebを参照
- 尿細管は伴走血管との間で物質の輸送を行う.1
- 尿細管は近位尿細管,ヘンレループ,遠位尿細管,集合管からなる.2
- 集合管は各ネフロンからの尿を集めて乳頭部へ流す.3
尿細管での物質輸送
▶tublar transport
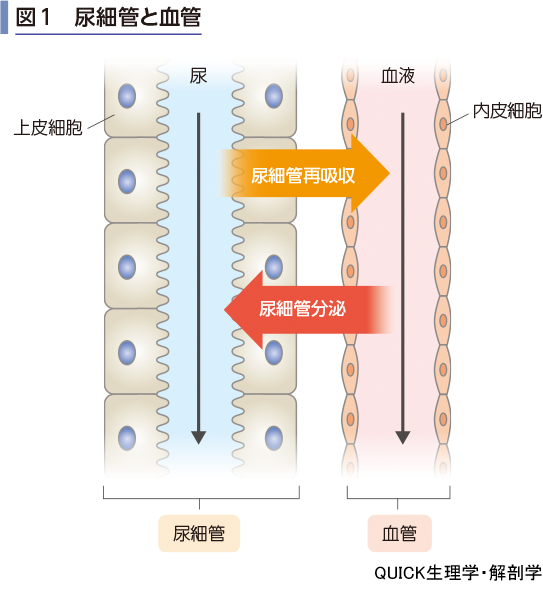
尿細管は尿中の物質を血液に戻したり,血液中の物質を尿中に排出したりする機能がある(図1).前者を尿細管再吸収,後者を尿細管分泌とよぶ.これは尿細管の周りには常に毛細血管が取り巻いていることから可能になる.尿細管を取り巻いている血管は,腎小体の輸出細動脈から枝分かれしている.つまり,腎小体を発した尿細管と血管はそのまま伴走しているのである.
尿細管の構造
▶structure of renal tubule
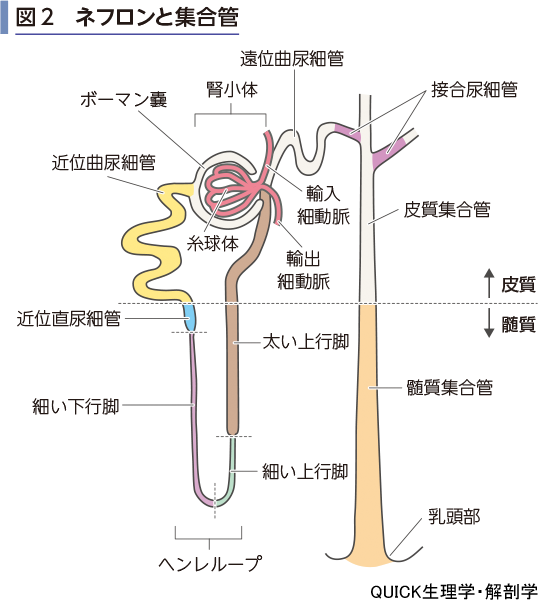
腎小体のボーマン嚢は尿細管に移行する(図2).これを近位曲尿細管とよぶ.近位曲尿細管は皮質内を走行し,髄質に降りてヘンレループへ移行する.近位曲尿細管の上皮細胞は,管腔側の細胞膜がブラシ様に毛羽立っており,刷子縁膜とよばれる.これは近位曲尿細管が尿から栄養素を再吸収するのが主な役割であり,再吸収のため尿と上皮の間の接触面積が広くなっている.
尿細管は髄質の中でヘンレループとなる.これは皮質側から髄質の奥までまっすぐ伸びる下行脚とそれが反転して皮質に戻る上行脚からなる.上行脚はさらに細い上行脚とその後の太い上行脚に分けられる.
皮質に戻った尿細管は遠位曲尿細管となり,自ら発した腎小体の輸入細動脈と輸出細動脈の間で糸球体と接している.さらに伸びて,近隣のネフロンの遠位尿細管と合流してゆく.この部分を接合尿細管といい,合流していく部分が集合管である.集合管はある程度のネフロンの合流を重ねたあと,髄質を貫いて乳頭部に開口する.
- 腎髄質に存在するのはどれか.(創作問題)
- a集合管b近位尿細管c腎小体d遠位尿細管e接合尿細管
- 解答と解説はWebを参照
- 栄養素の回収は近位尿細管で,100%行われる.1
- グルコースはSGLT2による二次性能動輸送で再吸収される.2
- アミノ酸の再吸収は,分子種により異なる輸送体を用いる.3
- ペプチド,タンパク質の再吸収も近位尿細管で行われる.4
原尿中の栄養素の回収
▶salvage of nutrients from primitive urine
原尿に含まれている栄養素は,グルコース,各種アミノ酸,ペプチド,アルブミンなど小さめのタンパク質分子である.これらは,近位曲尿細管を通過している間に100%完全に再吸収され,通常,尿中には出現しない.
グルコースの再吸収
▶reabsorption of glucose
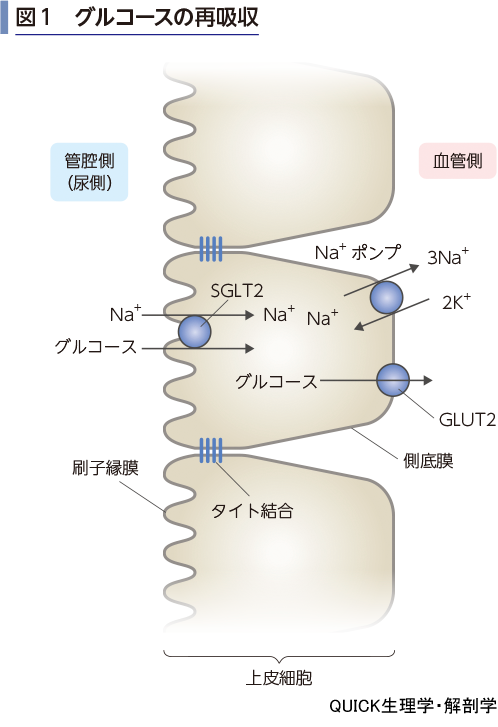
グルコースは刷子縁膜上のNa+-グルコース共輸送体(SGLT2)によって再吸収される(図1).この再吸収の原動力はNa+の濃度勾配である.原尿は血漿と同じ濃度のNa+を含むので,上皮細胞内のNa+濃度より10倍程度高い.したがって,濃度勾配の力によりNa+は細胞内に流入しようとする.この力を利用して,原尿のグルコースを同時に細胞内に引き込むのである.濃度勾配にしたがってNa+が流入すると,いずれは細胞内外の濃度が同じになり,濃度勾配が失われる.これを防ぐために,流入したNa+は側庭膜上のNa+ポンプにより能動的に血管側へ排出され,濃度勾配は維持される.いわゆる二次性能動輸送である.再吸収され高濃度になったグルコースは,側底膜のグルコース輸送体(GLUT2)により,濃度勾配にしたがって血管へ放出される.このグルコースの再吸収は近位尿細管において行われ,この機構が正常に働いている場合には,原尿のグルコースは100%再吸収される.
アミノ酸の再吸収
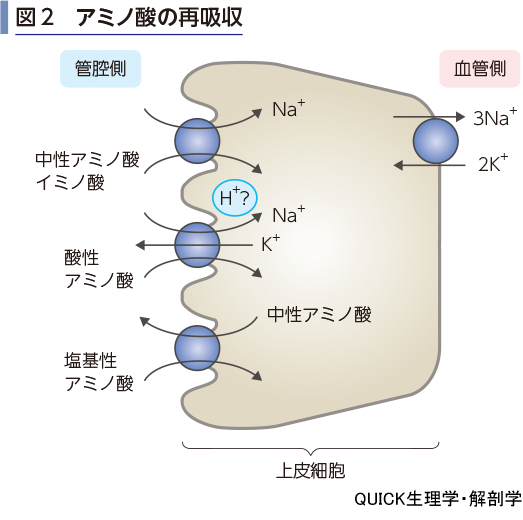
▶reabsorption of amino acids
尿細管上皮細胞でのNa+の濃度勾配は,アミノ酸の再吸収にも利用される.Na+-アミノ酸共輸送体により中性アミノ酸とイミノ酸は二次性能動輸送の形式で再吸収される(図2).酸性アミノ酸の場合もNa+濃度勾配に依存しているが,H+とK+の動きの関与が報告されている.塩基性アミノ酸は中性アミノ酸と交換する輸送体により再吸収される.
ペプチドおよびタンパク質の再吸収
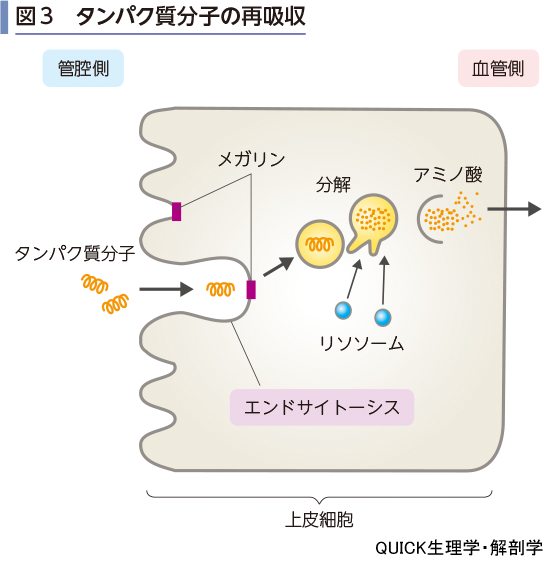
▶reabsorption of peptide and protein
原尿中のペプチドは刷子縁膜に存在するアミノペプチダーゼで分解され,アミノ酸となって再吸収される.また,ジペプチド,トリペプチドはH+-小分子ペプチド共輸送体(PEPT-2)により再吸収される.
糸球体濾過により血漿のアルブミン分画に属する低分子タンパク質は原尿中に出る.その量は1日あたり約4.5 gであるが,尿中排泄量は痕跡的であり,ほとんどが近位尿細管で再吸収されている.これらのタンパク質分子の再吸収は刷子縁膜でのエンドサイトーシスによって行われる(図3).上皮細胞内でリソソームと融合して,そのタンパク質分解酵素によりアミノ酸まで分解されて,血液中へ戻される.エンドサイトーシスを担う受容体はメガリン(LDL receptor related protein 2)とよばれる.
Tm制限性再吸収
Tm(transport maximum)とは,尿細管最大輸送量のことである.尿細管での再吸収や分泌のTmは,発現している輸送体の最大能力に依存して決まる.近位尿細管でのアミノ酸再吸収のTmは非常に大きく,原尿のアミノ酸がどれだけ増えても,すべて再吸収できる.一方,近位尿細管でのグルコース再吸収のTmはそれほど大きくなく,血糖値が200 mg/dLを超える原尿が近位尿細管を通過するとTmを超えているため再吸収しきれない.再吸収されなかったグルコースはそのまま膀胱に至り,尿糖として検出されることになる.これをオーバーフロー糖尿とよぶ.糖尿病は血糖値が高くなる病気であり,そのためオーバーフロー糖尿が生じ,尿中に糖が検出される.これが病名の由来であるが,病気の本質ではないことに注意すべきである.
- グルコースが尿細管を介して再吸収される際に最初に通過するのはどれか.(創作問題)
- a基底膜b側底膜c刷子縁膜dタイト結合eギャップ結合
- 解答と解説はWebを参照
- Na+の再吸収は,近位尿細管,ヘンレループの上行脚,遠位曲尿細管と皮質集合管で行われる.1
- Na+の再吸収は,主に二次性能動輸送による.1
- Cl-の再吸収は細胞を通過する経路と細胞間隙を通過する経路を用いる.2
- K+の再吸収は近位尿細管で受動的に,太い上行脚で能動的に行われ,皮質集合管主細胞では能動的な分泌が行われる.3
- アルドステロンはNa+とK+の尿への排泄量を決定する.4
- Ca2+の再吸収は上皮小体ホルモンの制御を受ける.5
Na+の再吸収
▶reabsorption of sodium ion
血漿にはさまざまな電解質が存在する.糸球体濾過では,そのすべてが濾過されることになるので,原尿におおむね血漿と同じ濃度,構成の電解質が存在する.その大半を占めるのは,Na+である.糸球体濾過されたNa+のおよそ1%程度が尿中に排泄されるので,99%の再吸収が尿細管を通過する間に行われることになる.Na+の再吸収の割合は,近位尿細管で70~75%,ヘンレループで20~25%,遠位尿細管および集合管で5~10%程度と見積もられる.
近位尿細管での再吸収
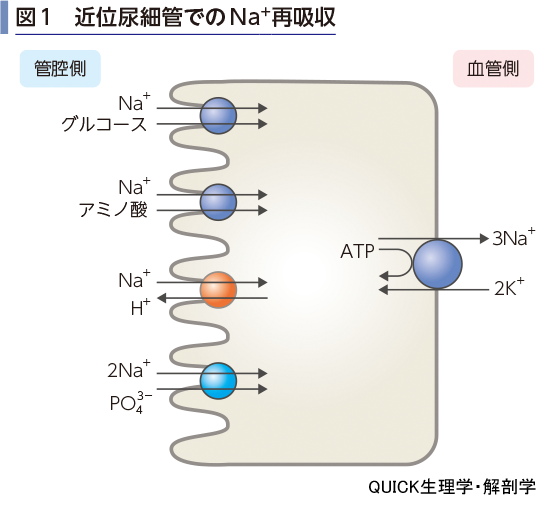
糸球体濾過されたNa+のほとんどを再吸収しなければならないので,近位尿細管では無条件に,ほぼ自動的な再吸収が行われる.グルコースやアミノ酸の回収で用いられているNa+ポンプによる二次性能動輸送は,Na+再吸収の主要な手段でもある(図1).さらに二次性能動輸送を用いる再吸収機構として,刷子縁膜(管腔側膜)に存在するNa+-H+交換系がある.これはNa+の濃度勾配を利用して上皮細胞内のH+を尿中に排出するしくみであるが,HCO32-の再吸収のしくみと関連している(第7章-08参照).原尿中のリン酸の再吸収を担うのはNa+依存性リン酸輸送体で,これもNa+濃度勾配を利用した二次性能動輸送である.
ヘンレループでの再吸収
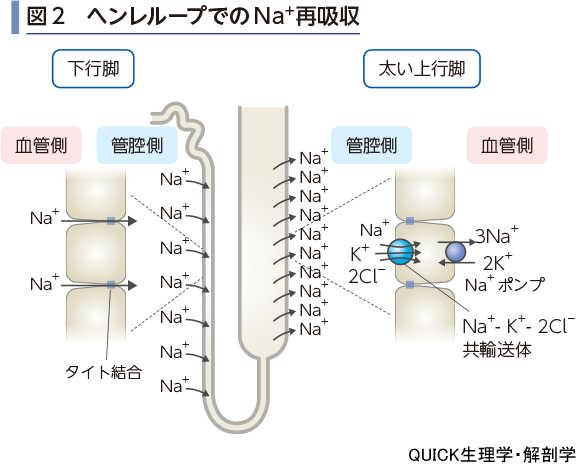
ヘンレループでのNa+の動きは複雑である(図2).下行脚では管腔側へのNa+の透過性が高いと考えられている.おそらくは上皮細胞間接着(タイト結合)を介する経細胞間隙経路であり,周囲の髄質が高Na+濃度であることから,Na+が尿中へ分泌される(しかし,動物種によっては膜透過性がないという報告もある).一方,太い上行脚では,Na+ポンプが豊富に存在し,管腔側膜にあるNa+-K+-2 Cl-共輸送体により,二次性能動輸送でNa+の再吸収が行われる.上行脚での能動的再吸収量が圧倒的なので,ヘンレループ全体での収支としてはNa+再吸収となる.
遠位曲尿細管と皮質内集合管での再吸収
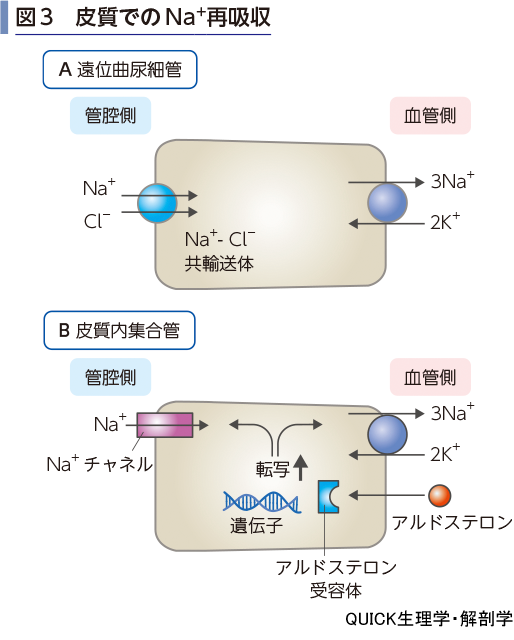
遠位曲尿細管では,管腔側膜のNa+-Cl-共輸送体を用いた二次性能動輸送がさかんである(図3A).さらに皮質内の集合管ではNa+チャネルを利用した二次性能動輸送が行われる(図3B).こうして尿を生成する最終段階でNa+の再吸収が能動的に行われるが,皮質内集合管でのNa+再吸収はアルドステロン(電解質コルチコイド)による制御を受ける.アルドステロンは皮質内集合管主細胞に作用して,Na+ポンプとNa+チャネルの転写,発現を促進することで,Na+の再吸収を増やす.したがって,最終的なNa+の尿への排泄量はアルドステロンが決定していることになる.
Cl-の再吸収
▶reabsorption of chloride ion
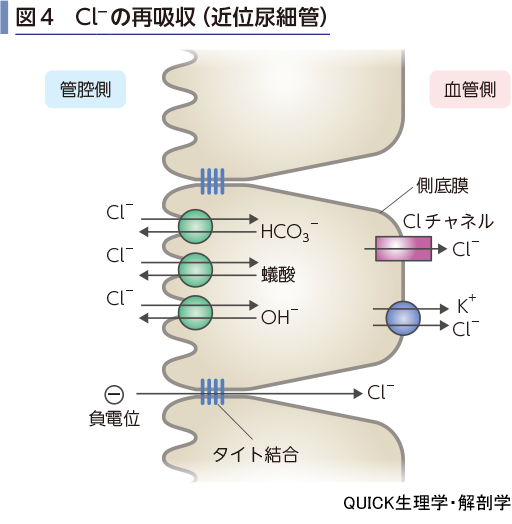
近位尿細管では刷子縁膜(管腔側膜)にあるCl-と他の陰イオン(HCO32-,OH-,蟻酸,オキザロ酸など)の交換により,Cl−は上皮細胞内に入り,側底膜のCl-チャネルやK+-Cl-共輸送体により間質・血管へ再吸収される(図4).また,近位尿細管内は負に帯電しているので,Cl-はタイト結合を経由して(経細胞間隙経路)間質へ移動する.
ヘンレループの下行脚ではCl-の管腔側への透過性もNa+と同様に高く,ある程度の分泌があると考えられる.一方,太い上行脚では,Na+-K+-2 Cl-共輸送体によるNa+の再吸収の際に同時に再吸収される.
遠位曲尿細管では,管腔側膜のNa+-Cl-共輸送体によりNa+とともに能動的に再吸収される.
K+の再吸収
▶reabsorption of potassium ion
あらゆる細胞の内液には,大量のK+が含まれている.その量は3,500 mEqと見積もられる.しかし,細胞外液および血漿中のK+は3.5~4.8 mEqと低い濃度で維持されている.この濃度の変動が大きいと,心臓や筋肉,脳細胞の興奮性に大きな影響を与える.食物のほとんどは細胞由来であるので,毎日一定程度(およそ100 mEq)のK+が吸収される.したがって,便に10 mEq排出されるとして,残りの90 mEqは腎臓から尿中へ排出されなければならない.糸球体濾過されたK+は尿細管で再吸収と分泌を受けて,K+濃度を維持している.
近位尿細管での再吸収
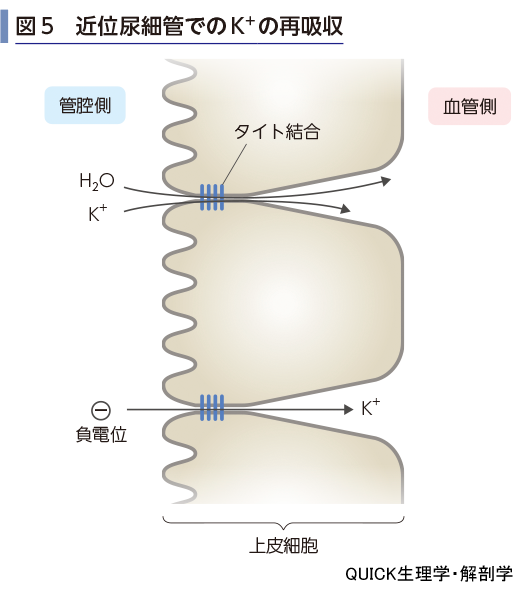
近位尿細管には積極的にK+を輸送する輸送体は存在しない.K+は主にタイト結合を通って再吸収される(図5).その原動力は,近位尿細管の前半では水の動き,後半では尿が負に帯電していることによる電気的駆動力である.
ヘンレループでの再吸収
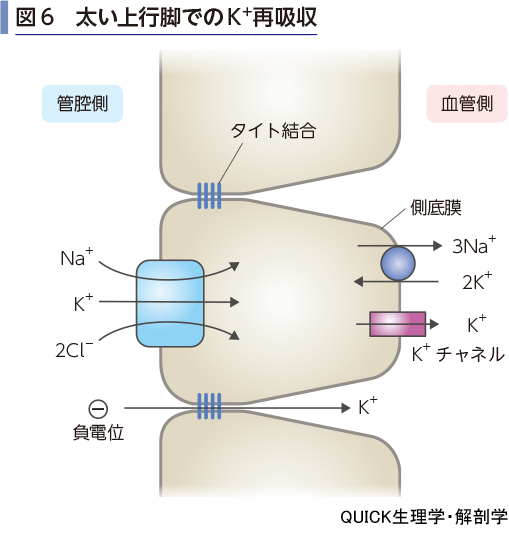
ヘンレループの下行脚ではNa+と同様に管腔側へのK+の分泌が行われる.太い上行脚ではNa+-K+-2 Cl-共輸送体による二次性能動輸送のしくみによって,K+の再吸収が行われる(図6).上皮細胞内に入ったK+は側底膜のK+チャネルを通って血液中へ移動する.また,電気的駆動力によりタイト結合を介する再吸収も報告されている.
皮質集合管での再吸収
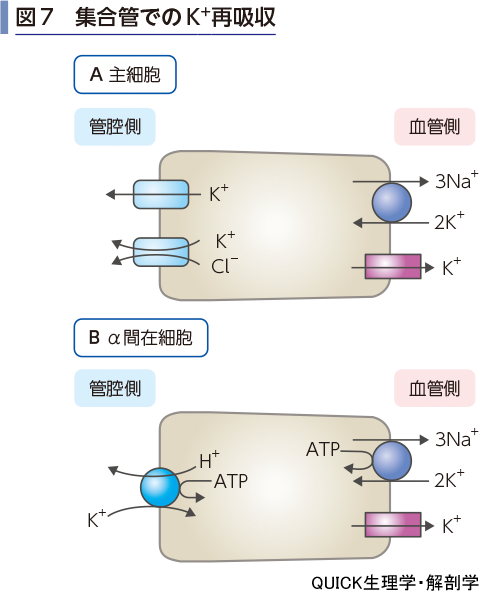
皮質集合管では2種類の上皮細胞が存在し,主細胞はK+チャネルによるK+分泌が行われる(図7A).一方,α間在細胞の管腔側膜にはH+-K+ATPaseが存在し,H+の能動的な分泌と引き換えにK+の再吸収が行われている(図7B).皮質集合管での輸送は血漿のK+濃度に敏感に反応すると考えられ,高K+濃度では主細胞でK+チャネルが増加して分泌促進,低K+濃度ではα間在細胞でH+-K+ATPaseが増加してK+の再吸収が促進される.
皮質集合管ではアルドステロンにより主細胞側底膜のNa+ポンプが増加するが,これによりK+の分泌は促進される.
尿細管のK+の分泌は尿流量依存性に増大することがわかっている.したがって,利尿薬使用や多飲の場合,通常より多くのK+が体外に排出される.
アルドステロンによる電解質輸送の調節
▶control of electrolyte transport by aldosterone
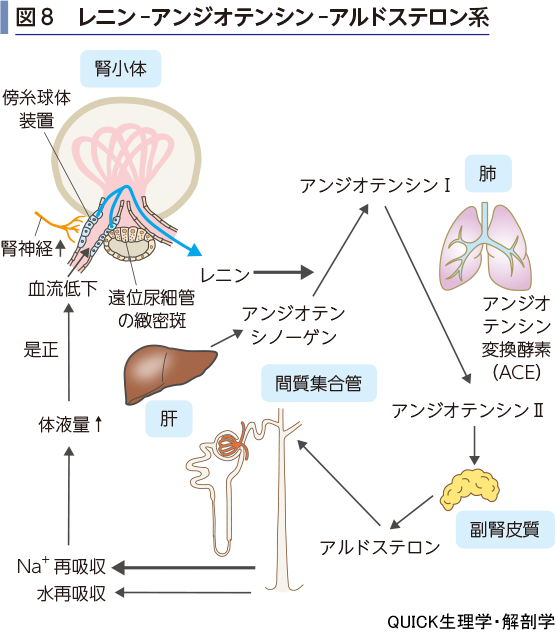
アルドステロン(電解質コルチコイド)は副腎皮質の球状層から分泌されるホルモンである(図8).腎臓の集合管主細胞に作用して,Na+の再吸収とK+の分泌を促進する.アルドステロンの分泌はレニン-アンジオテンシン-アルドステロン系によって制御されている.
レニンは輸入細動脈の傍糸球体装置から分泌されるタンパク質ホルモンである.レニン分泌は輸入細動脈に流れ込む血流量が低下することで促される.血流低下は全身の血圧低下や循環血液量の減少で引き起こされるので,これらの変化が分泌刺激となる.一方,尿中Na+濃度の低下は,遠位尿細管の緻密斑を介して傍糸球体装置を刺激しレニン分泌を促す.また,血圧低下は交感神経の興奮を引き起こすが,その枝は腎神経として傍糸球体装置に分布しており,交感神経刺激もレニン分泌を促す.レニンは高分子なので糸球体濾過されず,腎静脈に流れ出て,下大静脈に達する.下大静脈には肝臓からの血流も含まれており,肝臓で産生されたアンジオテンシノーゲンを多く含んでいる.レニンはアンジオテンシノーゲンに作用して,アンジオテンシンⅠを静脈血内で生成する.こうして生じたアンジオテンシンⅠは次に肺に流入する.肺の毛細血管の内皮細胞膜にはアンジオテンシン変換酵素(ACE)が豊富に存在するので,流れ込んだアンジオテンシンⅠはアンジオテンシンⅡに変換される.こうして大量のアンジオテンシンⅡは動脈へ流れ出す.アンジオテンシンⅡ自体は強力な血管収縮物質であるが,その他に副腎皮質球状層の細胞を刺激するので,アルドステロンの分泌が引き起こされる.
アルドステロンが集合管に作用すると,Na+の再吸収が促進される.そうすると,血漿のNa+が増えるので,血漿浸透圧が上がる.そこで浸透圧を維持するために尿細管の各部分での水の再吸収が促進され,結果的に循環血液量は増加することになる.したがって,全身の血圧は上昇することになる.こうして,レニン分泌のきっかけとなった変化は是正されたことになる.
高血圧治療薬
現在臨床使用されている高血圧治療薬のうち,かなりのものはレニン-アンジオテンシン-アルドステロン系の働きを抑えるものである.アンジオテンシン変換酵素阻害薬はアンジオテンシンⅡの産生を抑え,アンジオテンシンⅡ受容体阻害薬はアンジオテンシンⅡが血管を収縮させることを抑える.また,抗アルドステロン薬も導入され,臨床応用されている.これらのことは,正常の血圧制御にレニン-アンジオテンシン-アルドステロン系がきわめて重要な役割をもっていることの反映である.
Ca2+, PO43-の再吸収
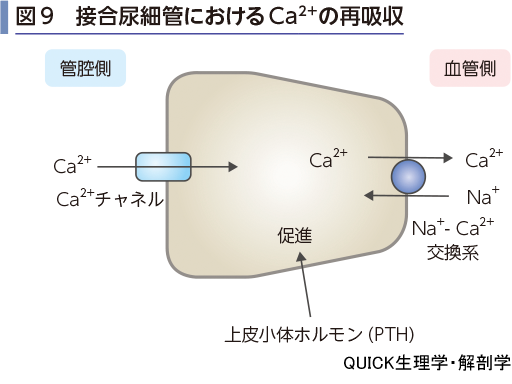
▶reabsorption of calcium ion and phosphate ion
Ca2+は糸球体濾過された分の95%以上が再吸収される.近位尿細管での再吸収は60~70%であるが,機序はまだ明らかではない.ヘンレループでは上行脚でCa2+チャネルを介した再吸収が行われ,糸球体濾過された分の20~25%になる.遠位尿細管および集合管では10~15%の再吸収があり,特に接合尿細管ではCa2+チャネルからNa+-Ca2+交換系を介する再吸収が行われている(図9).上皮小体ホルモン(PTH),この再吸収過程を促進する.
PO43-は糸球体濾過された分の80%程度が尿細管で再吸収される.再吸収の大部分は近位尿細管で行われる.刷子縁膜に存在するNa+とPO43-との共輸送体(NaPiⅡa)により,細胞内に取り込まれる(図1).しかし,側底膜での再吸収ルートはまだ不明である.NaPiⅡaは上皮小体ホルモンにより抑制される.
- 尿細管でのNa+の輸送にかかわるのはどれか.(創作問題)
- ①タイト結合
- ②Na+ポンプ
- ③プロトンポンプ
- ④水チャネル
- a①,③,④があてはまるb①,②があてはまるc②,③があてはまるd④のみあてはまるe①~④すべてがあてはまる
- 解答と解説はWebを参照