第Ⅳ部 ニューラルネットワークの基礎
第11章 脳とニューロン
廣瀬 明
(東京大学大学院工学系研究科電気系工学専攻/バイオエンジニアリング専攻)
 脳とは何か:モノとしての側面
脳とは何か:モノとしての側面
パーソナルコンピュータ(personal computer:PC)は,詳細に設計された中央処理装置(central processing unit:CPU)や誤りのない回路基板を使って構築され,精密な計算を行っている.一方,脳は極めて大雑把な構造が遺伝子に書き込まれているのみであり,その意味で詳細に作りこまれているわけではない.それなのになぜ,人の顔を瞬時に認識するといった,高速コンピュータでも苦手な処理を脳は簡単にやってのけるのか.実は,コンピュータと脳とでは,情報処理の方式の原理が異なっている.本章と次章ではこの点を概観することにする.まず本章では,モノの側面を見てみよう.上述のように,脳は「作り込まれていない」構造をとっている.次章の内容とあわせて考えると,そのことが,脳の柔軟性と深く結びついていることがわかる.また本章第6節では,生命科学やバイオ分野全般における物理や数理の重要性も指摘する.
 脳とコンピュータの外見的・機能的な違い
脳とコンピュータの外見的・機能的な違い
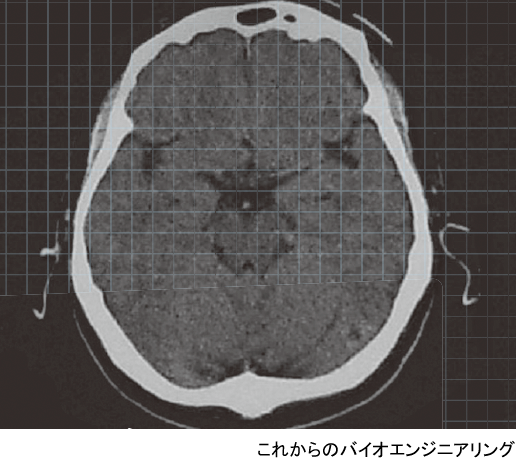
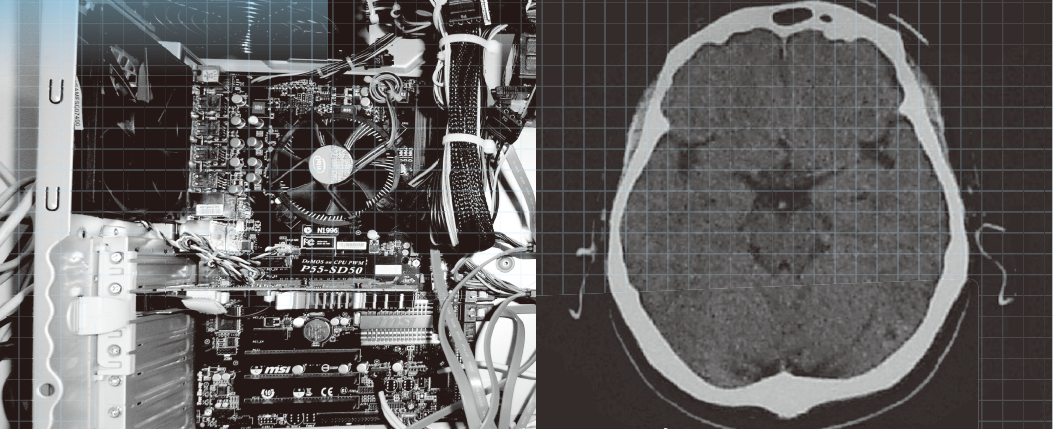
冒頭左画像は,PC の中をのぞいてみた写真である.PC にはCPU,主メモリとなる半導体メモリ,外部記憶装置のハードディスクドライブ(HDD)や半導体ディスク(SSD)などの部分があることがわかる.それらが回路基板やケーブルによって接続され,全体としてPC を構成している.
一方,右は脳内の構造をMRI(magnetic resonance imaging:核磁気共鳴イメージング)によって得た画像である.眼球より少し上の水平面で見たものである.大きな面積を占める灰色の部分が脳であり,白い頭蓋の中に納まっている.中心付近には視床などの間脳や脳室があり,生命活動の基本を支えている.その外側の頭蓋に近いところは大脳皮質と呼ばれ,ヒトの高度な情報処理機能はここを中心に行われている.大脳皮質は2次元的な広がりを持ち,それが折りたたまれるようにして頭蓋内に納められている.いわゆる,脳の
脳とPC ではこのような構造の相異があるにもかかわらず,脳はどうして高度な情報処理を行うことができるのだろうか.またその処理の高度さも,コンピュータと脳とではその得意・不得意が異なる.コンピュータは桁数の多い計算も難なくこなし,論理的な処理が得意である.しかし,人の顔を瞬時に判別したり,会話の微妙なニュアンスを感じ取ったりすることは苦手である.これらの点は,脳と対照的である.
 ニューロン
ニューロン
脳を構成する主役の素子は,ニューロン(神経細胞)である.図11.1 A は,ニューロンを着色して光学顕微鏡で見たものである.細胞体の大きさは数 μm~数百 μm(マイクロメートル = 10−6 m)である.コンピュータのトランジスタの大きさが10 nm(ナノメートル = 10−9 m)程度なのに比べて千~十万倍と大変大きい.細胞体からは入力信号を集める
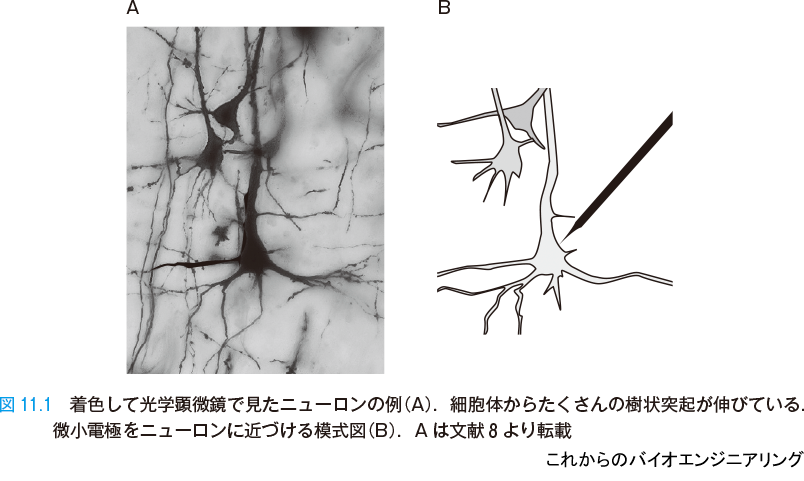
図11.1 B は,ニューロンに微小電極を近づける様子を示す模式図である.微小電極は,極細のガラス・ピペットである.中に生理食塩水を入れると,導電性を持たせることができる.また外部とはガラスで絶縁され,ちょうど先端を小さな電極として使うことができる.微小電極を細胞膜表面に当ててオシロスコープで観測すれば,膜の外面の電位を時刻に対する変化として計測できる[1].そのとき,接地電極は脳切片の端などの遠方に置いて,基準電位にする.微小電極を細胞に突き刺してわずかに先端を内部に入れれば,細胞内の電位を計測できる.細胞がパルス電圧を出力する(発火する)ことは,これで計測できる.また近年,膜電位に応じて光の反射率が変化する色素を利用するオプティカル・レコーディングなども用いられ,応答を2次元的に広範囲に計測することも広く行われている.
 ネットワーク,層構造,コラム構造
ネットワーク,層構造,コラム構造
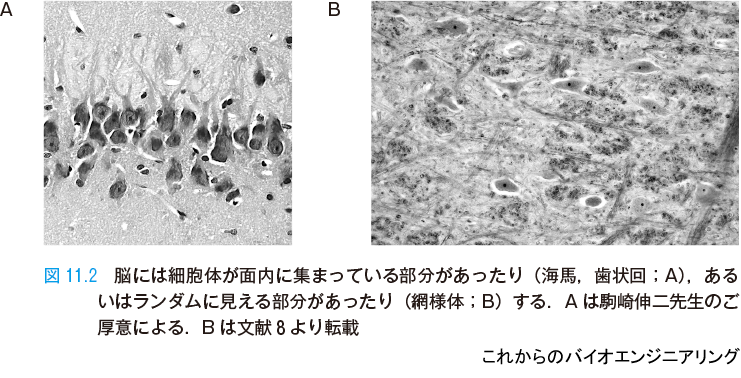
ニューロンは軸索を伸ばしてネットワークを作り,他のニューロンの入力部である樹状突起や細胞体に出力信号を届けている.図11.2 は,たくさんのニューロンがどのように配列しているかを示す例である.図11.2 A ではニューロンは面内に細胞体が集まっていて,層構造をもっている.この層から上下に樹状突起や軸索が伸びている.面内に伸びるものもある.一方,図11.2 B のようにランダムな部分もある.
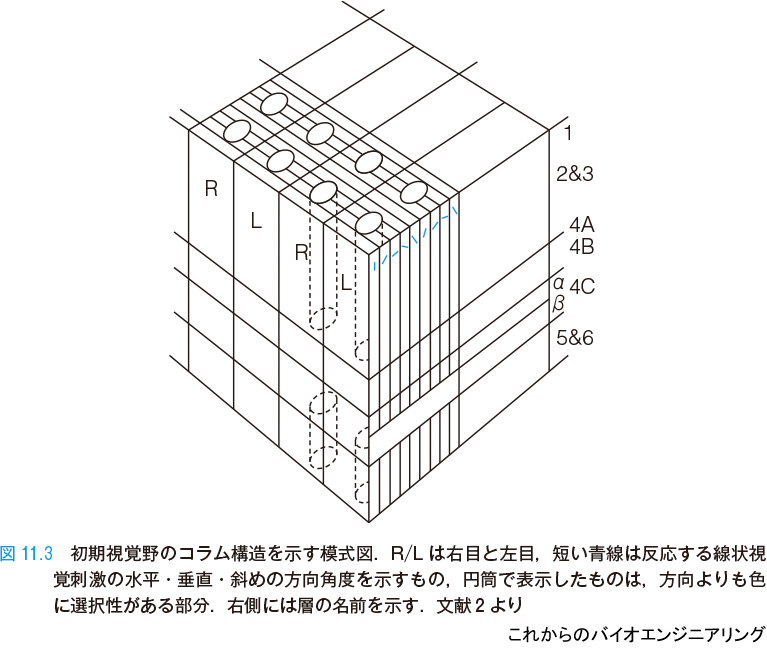
ヒトらしい機能を実現している大脳皮質の新皮質は,2 mm ほどの厚さの中に6層をなしている.図11.3 のように頭蓋に近い第1層から,2 mm ぐらいの深さにある第6層までの層構造をもち,その面積は広く2,200 cm2 程度あり*1,脳の表面に皺を作りながら折りたたまれるように格納されている.ニューロンが作る層構造は,その処理方式と密接な関係がある.人工知能(artificial intelligence:AI)では,ディープラーニング(深層学習)などを含む層状ニューラルネットワークが最も一般的な構造であるが,その発想の元がこの皮質の層構造である.
大脳皮質は,層を貫く向きにコラム構造をもつところもある.樹状突起がおもに層を直交する方向に伸びて深さ方向に信号をやり取りする.
 トポグラフィック・マッピング
トポグラフィック・マッピング
上記の構造は,信号処理機能と深い関係を持つ.図11.3 に示す初期視覚野(後頭部の皮質)では,右目からくる信号を処理するところ(R)と,左目からくる信号を処理するところ(L)が交互に配置されている.また水平な視覚刺激(横棒をみせる)に反応するニューロン,少し右肩下がりに傾いた刺激に反応するニューロン,もう少し方向が右肩下がりの刺激に反応するニューロン…というように,視覚刺激がもつ方向に反応するニューロンが,刺激角度の近さ遠さをそのまま反映するように整然と,少し重なり合いながら,並んでいる.このように,反応するニューロンの位置に意味がある信号の伝達や処理を,トポグラフィック・マッピング(topographic mapping:地形的射像)と呼ぶ.
レチノトピー(retinotopy:網膜+位置の意味)と呼ばれるものも,その1つである.網膜(レチナ:retina)に作られた像はそのまま像の形を保つように初期視覚野に投影される.網膜に映った形が,少し
それは,脳全体の役割分担にも見られる.運動生成に関係する脳のどの部分が体のどの部位を制御しているかを,模式的に示す(図11.4).脳を前面から見たときの頭頂から側頭までを表している.頭頂は足や体幹の制御を担当している.手が大きく表現されているが,複雑な動きを実現するために脳の大きな面積を占有している.その他,唇や顎,舌といった発話に関係する部分にも大きな面積が使われている.このように体の部位の制御に使われる皮質の部分はそれぞれ局在している.しかもその分布は体のつながり具合の構造をそのまま反映したものになっている.すなわち,体の部位が隣り合えば,皮質上の部位も隣り合っている.このような対応(マッピング)も,トポグラフィック・マッピングである*2.
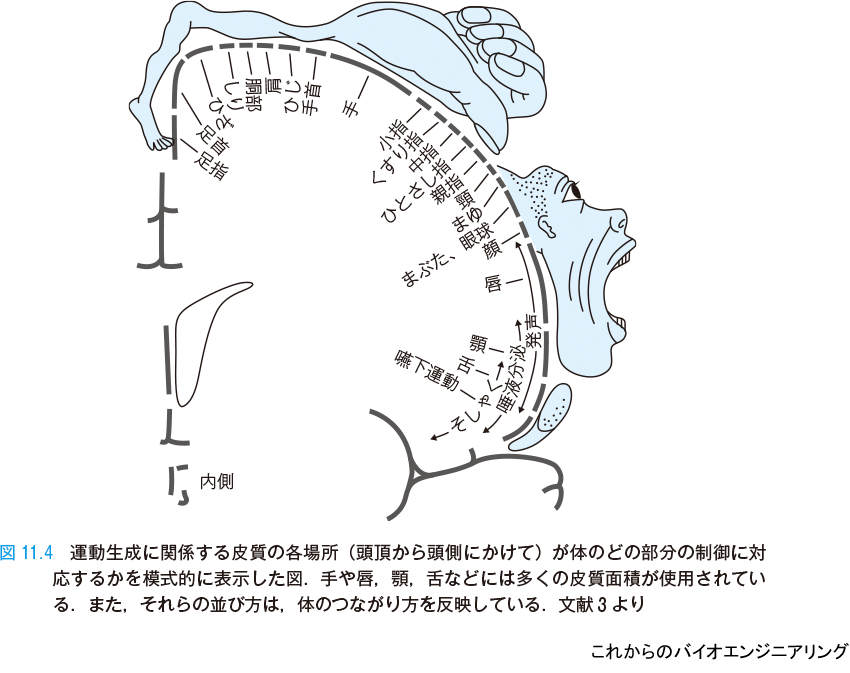
このように,トポグラフィック・マッピングは脳の大きな特徴である.コンピュータの構造に詳しい読者やプログラミングを行ったことのある読者は,コンピュータではこうなってはいないことを感じるのではないだろうか.コンピュータでは,すべての情報を限られた数の局在するCPU が処理する.メモリに情報を格納する際にも「隣り合う情報は隣り合うところに」ということを殊更には行っていない.コンピュータのメモリのやり方を線形番地,脳のやり方を内容番地あるいは連想番地と呼ぶこともある.脳では処理される情報と,処理される場所には深い関係がある.
もっとも,体のマッピングについては次のように考える読者もいるだろう.1つの細胞から発生した個体が細胞増殖で体を形作るのだから,体の各部分とそれを制御する脳の各部分をつなぐつなぎ方は,隣り合う者同士が離れないようにすることは,自然の成り行きだ.確かにその通りだろう.しかし,はじめに挙げた「似た視覚刺激は隣り合う皮質の個所で処理される」というマッピングはどうだろう.視覚刺激の方向(角度)は情報自体がもっている性質であって,情報の近さ遠さが脳の処理部分の近さ遠さを決めることは,自明ではないだろう.これは,12章で述べる脳の自己組織化によって実現されるものである.
このような情報の近さ遠さを保つマッピング,すなわちトポグラフィック・マッピングは,脳の柔軟性に大きな役割を果たしている.脳では毎日,相当の数のニューロンが死んでいる.しかし,隣り合うニューロンが類似の情報を扱っていたならば,隣りのニューロンが死んだニューロンの代理として活躍することも不可能ではない.線形番地のコンピュータではこうはいかない.コンピュータの中で毎日,相当の数のトランジスタが死んでいたら,コンピュータは機能しないだろう.
 信号は何か
信号は何か
神経情報を表す信号は何か.4および6章で概略を見たように,細胞膜の内側と外側には電位差がある.これを膜電位と呼ぶ。ニューロンが反応する,すなわち発火するとはニューロンが膜電位の電圧パルスを出すことである.この意味で信号は電気的なもの(電圧,電流)である.電位を頭皮の電極で計測すればEEG を得るし,電流を頭外の磁場として計測すればMEG を得る(10 章参照).もっとも,軸索の先端で次のニューロンに信号を伝えるところは,化学的な物質の放出と受容によっている.しかし信号の基本は電気であるといってよい.
電位の発生の機構を説明する(図11.5)[1].灰色の部分が細胞膜で,その上側は細胞の外,下側は細胞の中を表している.細胞膜は,化学的にはいわゆる半透膜である.それによって隔てられた2つの領域に存在するイオンの濃度が決まると,その間に発生する電位も決まる.化学的ポテンシャルと電気的ポテンシャルが釣り合うように平衡状態が生成される.そのため,膜間の電位を議論するためにはそれぞれのイオン濃度を議論する必要がある.
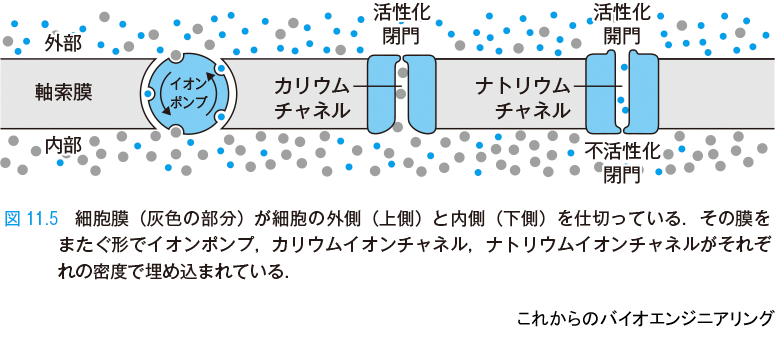
通常,細胞外にはナトリウムイオン濃度が高く,細胞内にはカリウムイオン濃度が高い.細胞膜には膜をまたぐ形でイオンポンプが埋め込まれている.イオンポンプは細胞内のナトリウムイオンを放出し,外のカリウムイオンを取り込んで,常に一定のイオン濃度差が生じるように動作している.その結果,例えばヒトの脳細胞では静止電位*3 はおよそ −60 mV である.
細胞膜にはカリウムチャネルやナトリウムチャネルといった特定のイオンのみを通したり通さなかったりする通路(チャネル)も埋め込まれている.これらのチャネルの分子は膜電位を感じ取って自らの分子形状を変え,イオンを通したり堰き止めたりする.そのイオン電流の通しやすさ(コンダクタンス)は膜電位に対して非線形に変化する.そのため,膜電位がある一定の値(
ニューロンの軸索はパルスを運び,シナプスを介して信号を他のニューロンの樹状突起や細胞体に伝える.たくさんの他のニューロンから送られてきた信号(= 電圧 ≈ 膜をはさんだ電荷)が細胞膜という共通のコンデンサで合算されて,膜電位を脱分極の方向にゆさぶる.それが細胞体でのパルス生成を引き起こす(すなわち発火).パルスの尖頭値はおよそ +40 mV である.
細胞体で発生したパルスが軸索を伝搬する様子を示す模式図を示す(図11.6)[1].チャネルの非線形性によって,パルスはその形を崩すことなく安定して伝搬する.A はパルス発生から 2 ms 後,B は 6 ms 後,そしてC は 10 ms 後の様子である.パルスの伝搬速度はおよそ 1 m/s 程度であるが,速い場合には数十 m/s になる.それは軸索の太さなどによって決まり,太いほど速くなる.
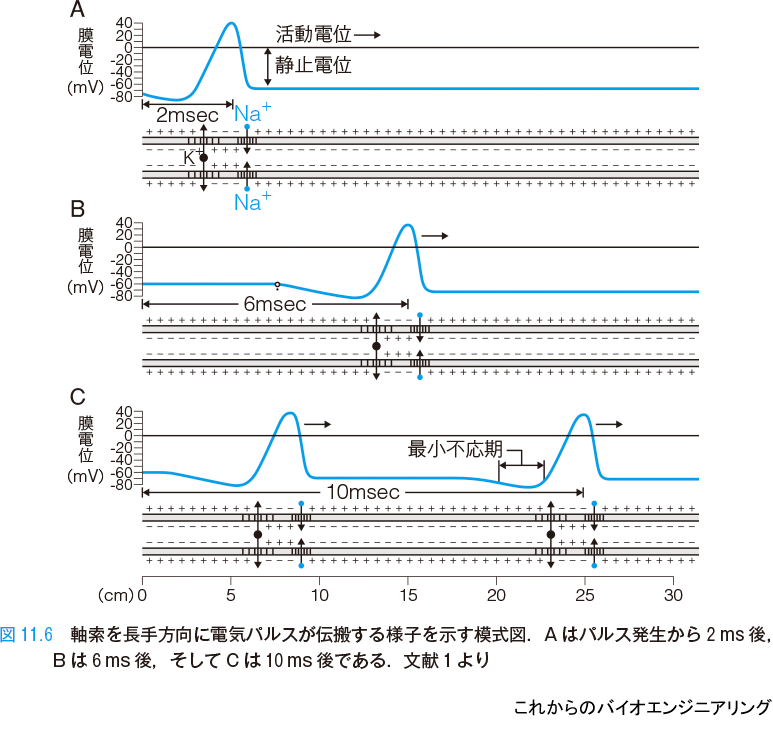
パルスの直後には不応期と呼ばれるパルス発生が不可能な期間がある.これはイオンチャネルが元に戻るのに必要な時間である.そのため,単位時間あたりに発生可能なパルスの密度には上限が生じる.パルスの頻度を周波数で表すとき,上限は 100Hz 程度になる.入力されるパルスの合計頻度がどんなに大きくても,発火のパルス頻度はこれを上回ることはなく,出力パルス密度は入力に対して飽和する.また,入力パルス頻度がある程度以上大きくならないと膜電位に影響を与えない場合も多い.そして影響を与え始めるパルス頻度の閾値がニューロンによってそれぞれ異なることもある.
 ホジキン・ハクスレー方程式
ホジキン・ハクスレー方程式
1940 年代後半,ケンブリッジ大学のホジキン(Alan Lloyd Hodgkin)とハクスレー(Andrew Fielding Huxley)は微小電極の直列抵抗を大幅に減らすなどの工夫を凝らし,膜間の電位を強制的に所望の電位に固定する膜電位固定法と名付ける計測方法を開発した.そして膜電位に対する膜電流の時間変化をさまざまな条件で詳細に計測することに成功した.1952 年,一連の実験に基づいて次のような1組の定量的な方程式を提案した[5].彼らはその後,この研究でノーベル賞を受賞することになる.
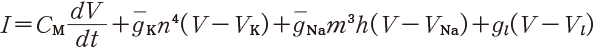 (11.1)
(11.1)
ただし,
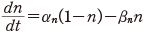 (11.2)
(11.2)
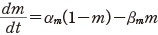 (11.3)
(11.3)
 (11.4)
(11.4)
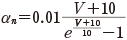 (11.5)
(11.5)
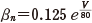 (11.6)
(11.6)
 (11.7)
(11.7)
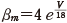 (11.8)
(11.8)
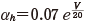 (11.9)
(11.9)
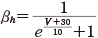 (11.10)
(11.10)
その発想と解析では,物理と数理が極めて重要な役割を演じている.これらの方程式は,次のように考え出された.ホジキンとハクスレーはまず,細胞膜の電気的な特性を図11.7 A のようにモデル化した.細胞膜は脂質二重膜で基本的には絶縁体であり,それをコンデンサとして表す.その単位面積あたりの容量を CM[μF / cm2] とする.微小電極を使い,膜電位固定法で細胞の内側と外側の間の電圧を所望の電圧にしたときの,微小電極を流れる電流の波形を計測した(図11.7 B)*4.この図で縦軸は,カリウムに対する単位面積あたりのコンダクタンスであり電流の大きさに対応する.
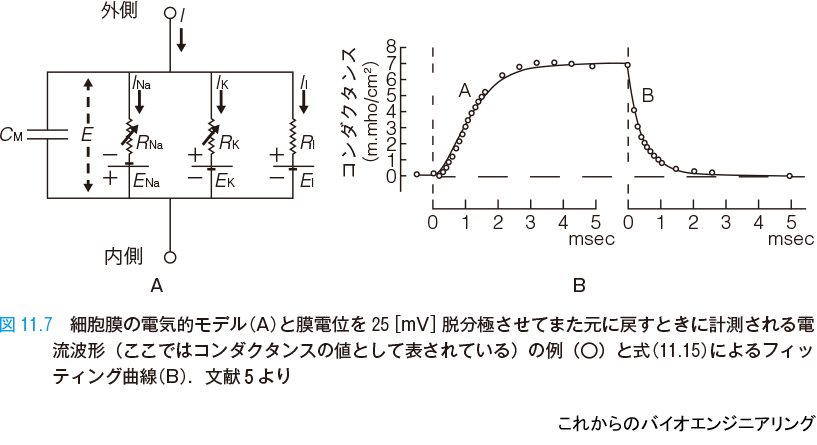
領域A は膜電位固定法でステップ関数的に 25[mV] だけ脱分極させたときの電流の立ち上がり波形であり,領域B は電位を元に戻して再分極させたときの立ち下がり波形である.図中○印が実測点である.ここから静止電位でのカリウムのコンダクタンス gK0 と電位を 25[mV] 脱分極させて十分に時間がたったときのコンダクタンス gK∞ ,および立ち上がりや立ち下がりの時定数 τn が見積もられた(表11.1).

ここで問題になるのは,領域A の立ち上がり初期が緩やかであることである.もし1次の変化,すなわち (1−e−t) の形ならば,初期は急峻に立ち上がらなければならない.立ち上がりの鈍り具合から,これは3次か4次の変化と見積もられた.どちらかというと4次とするとよく合致し,すなわち彼らは立ち上がりは (1−e−t)4 ,立ち下がりは (e−t)4=e−4tの形の変化であると考えた.
そのため,式(11.1)でカリウムのコンダクタンスは 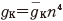 の形であるとし
た.ただし n は式(11.2)の中の n であり,再掲すれば
の形であるとし
た.ただし n は式(11.2)の中の n であり,再掲すれば 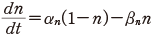 で
ある.この一般的な式の意味は, n (コンダクタンス値相当)がこの上限からど
のぐらいの割合であるかを表す変数で0から1の間の値をとること,それが αn と βn の時間変化率でその開閉の割合を変えること,である.また n4 の4乗は,そのようなゲートが4つあって全体として積の形で効くような機構をもつことを示唆する.
で
ある.この一般的な式の意味は, n (コンダクタンス値相当)がこの上限からど
のぐらいの割合であるかを表す変数で0から1の間の値をとること,それが αn と βn の時間変化率でその開閉の割合を変えること,である.また n4 の4乗は,そのようなゲートが4つあって全体として積の形で効くような機構をもつことを示唆する.
この αn と βn も実験から次のようにその値が得られる.静止電位の状態の n を n0 とすると,定常状態を考えて次の関係を得る.

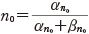 (11.11)
(11.11)
また,同様に膜電位をステップ的に変化させれば,その立ち上がりの様子と,十分に時間がたったときの αn∞ や βn∞ ,n の値について次の関係を得る.
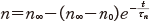 (11.12)
(11.12)
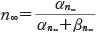 (11.13)
(11.13)
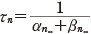 (11.14)
(11.14)
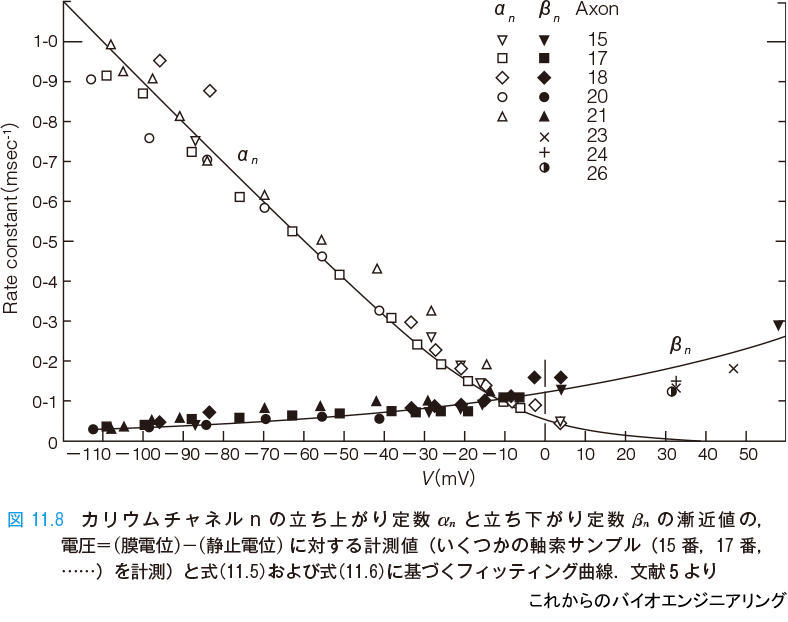
実験値は表11.1 であったことから,さまざまなクランプ電圧 V に対して電圧の関数として αn∞(V)と βn∞(V)(図中では αn とβn と表記)を得ることができる(図11.8).そして,式(11.5)と式(11.6)の具体的な定数を決めることができる*5.
そして,図11.7 B の実験をうまく表す曲線として,(1–e–t)4 の形に基づいて次のフィッティング曲線を得る.
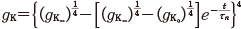 (11.15)
(11.15)
ナトリウムについても類似の考えで定数を決めてゆく.その結果,全体として式(11.1)が得られる.
(中略)
文献
- 「図説 脳」(塚田裕三/編),日経サイエンス,1983
- Margaret S. Livingstone and David H. Hubel, “Anatomy and physiology of a color system in the primate visual cortex,” The Journal of Neuroscience, 4, 309–356, 1984
- Theodore B. Rasmussen and Wilder Penfield, “The human sensorimotor cortex as studied by electrical stimulation,” Federation Proceedings Journal, 6 452–460, 1947
- Klaus Obermayer and Gary G. Blasdel, “Geometry of orientation and ocular dominance columns in monkey striate cortex,” The Journal of Neuroscience, 13, 4114–4129, 1993
- Alan L. Hodgkin and Andrew F. Huxley, “A quantitative description of membrane current and its application to conduction and excitation in nerve,” Journal of Physiology, 117, 500–544, 1952
- Erwin Neher and Bert Sakmann, “Single-channel currents recorded from membrane of denervated frog muscle fibres,” Nature, 260, 799–802, 1976
- 徳永太,中島龍一,シリーズ『若手の会インタビュー』日生誌,66,92–95,2004
- 「バーチャルスライド組織学」(駒崎伸二/著),羊土社,2020