
第16章 生物に情報記録マシンを仕込む—私たちの次のX
必要が発明の母であるならば,空想は発明の父であろう(ガリレオ・ガリレイが「懐疑は発明の父」であると残したと言われるが,こちらは「友人」くらいでよい気がする).かつてのドラえもんの主題歌にあったように「こんなこといいな,できたらいいな」と夢を見て,仲間達とそれをどうやったら実現できるだろうと積極的に考えて,挑戦してみることはとても楽しい.今の生物学の本質的な限界は何だろうか?生物学が明らかにしたいのにできていないことは何だろうか? それを突破するためにはどんなことができるといいだろうか?それはどうやったらできるだろうか? ──本書の最終章では,私たちのグループが推し進めているDNAイベントレコーディングのビジョンを紹介したい.ぜひこれまでの章を思い出しながら読んでみて欲しい.きっとよりおもしろいと思ってもらえるはずである.
はじめに
どのような生命現象も複雑であり,これを完全に捉えきることはできない.生物学におけるすべての発見は事象の側面であり,私たちはそれらをできるだけ多く集めて本質を抽象化しながら知識を構築していくことしかできない.だからこそ,対象を観察するための技術がとても大切である.一方で,「観察手法が今はこれしかないので,しかたがない」という考え方は私の研究グループはしない.また,「こういうトリックを考えるとこういう観察ができるからおもしろい」という考え方もたまにはいいが,常に現状に囚われず,「何を知りたいか,何がわかることが重要か」を想像するべきだと考える.それが新しい観察手法の必要性に至るのであれば,さらにそれが挑戦に値するか検討し,そのうえで挑む.いま私たちは細胞がその機能を動的に変化させながら複雑な組織を形成・維持するプロセスにおいて一つひとつの細胞すべての内外で何が起きているのか観察できる本質的に「強い」方法をつくり,一つの受精卵から哺乳動物(マウス)が全身を構成する過程の地図を細胞レベルで明らかにしたいと考えている.
DNAイベントレコーディング
動物の発生,また個体の恒常性維持と不全に対する応答においてはさまざまな種類の細胞が協奏し動的にその機能を変化させながら進展する.さまざまな動物において異なる組織や細胞に焦点を合わせてそれらがどうふるまうかについてはすでに多くの観察がされており,私たちに素晴らしい視点を与える.一方で,一見当たり前のことのように聞こえるが自然科学は目の前に存在する対象を観察することしか許さない.地球上にあまねく存在する多様な事象を観察することはできても,すでに絶滅した生物の発生過程を調べることはできない.シングルセルトランスクリプトーム技術(scRNA-seq)によって組織をどのような細胞が構成し,それぞれがどのような分子プロファイルをもつか解析できるようになったが,これも観察時点において細胞を破壊するので,それらの細胞が過去にどのような状態にあったのか,観察のために介入して破壊することがなければそれらがどのような未来を辿ったのか知る由もない.マウスの腫瘍の状態を調べるために凍結切片を観察することもできるだろうが,それを見てもその過去の状態や未来の状態はわからない.
「この箱を開けた時点で時間発展がわからなくなる問題は量子状態の観察のようである」とはoptimal transport(最適輸送理論)による細胞動態の推定手法1)を2019年に発表した数学者のGeoffrey Schiebingerの言葉であるが,言い得て妙である.無論,経時的なイメージングによって細胞や分子の動態を観察する手法はあるが(カルシウムイメージングなど),これらは顕微観察技術が対象にできる範囲の小さなシステムの観察しか許さない.私たちはまだ多細胞が構成する生命システムについて高い解像度の大規模情報を得ようとする場合は,違う風景のスナップショットを撮影して知識を抽出しようとすることしかできない.
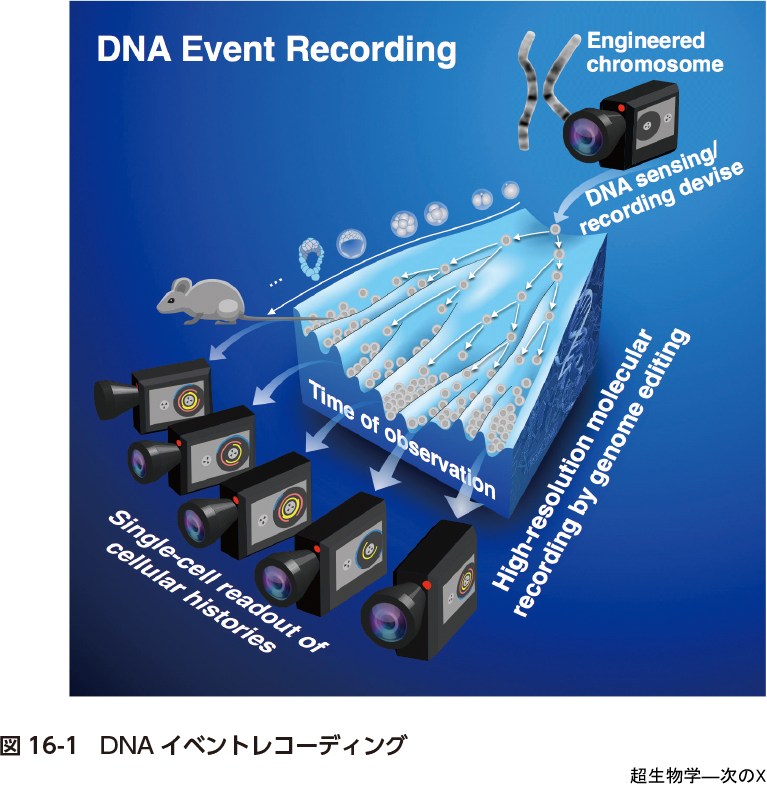
この生物学において当たり前な観察の制約を乗り越えるために価値のある挑戦は「高度なDNAイベントレコーダー」の開発であると私たちは考えている(図16-1).端的にいうとビデオカメラのような自律的な観察記録システムを人工遺伝子回路として細胞に搭載し,これが細胞の内外で起こるさまざまなイベントを細胞に搭載された人工的なDNAに(磁気テープのように)経時的に書き込んでいくようにしようというものである.細胞が分裂するたびに情報が記録されたDNAテープを娘細胞に継承するので,例えばこれが仕掛けられたマウスの受精卵から得られた成獣マウスはその細胞一つひとつが過去の状態ヒストリー情報をDNAテープにもつことになる.したがって,依然として観察時点において対象を破壊せずに情報を得る方法はないが,成獣を構成するすべての細胞についてDNAテープをシークエンシングしてそこから情報を再構築できれば動物が発生する過程を丸ごと捉えられる.またその情報の解像度は細胞に搭載するDNAイベントレコーダー,DNAシークエンシング,情報再構築手法の性能しだいということになる.
DNAイベントレコーディングの構成要素
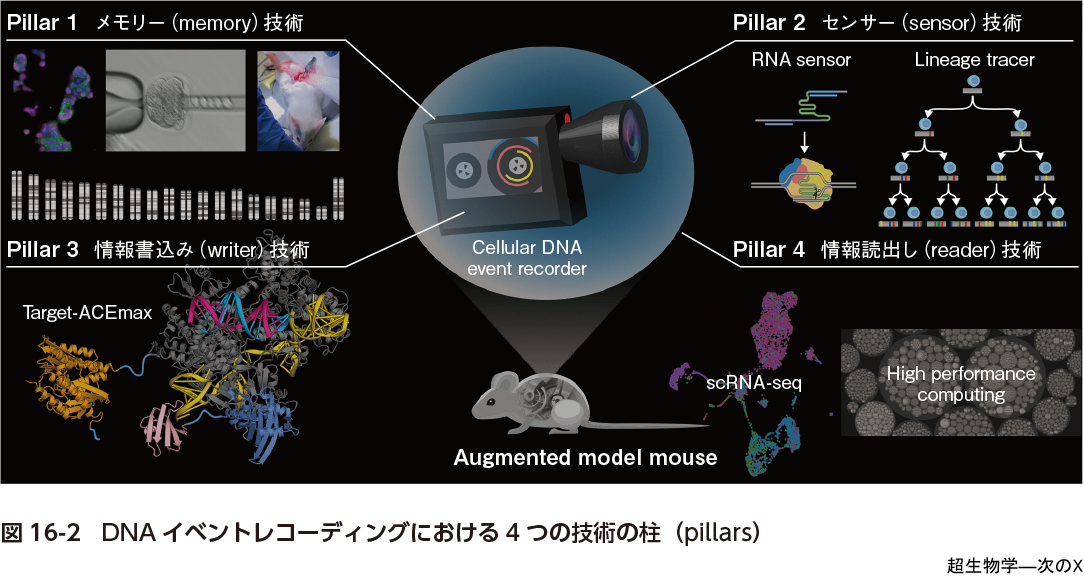
DNAイベントレコーダーが生物学の「箱を開けたら時間発展がわからなくなる」問題を解決する唯一の方法であるとは決めつけないが,私たちはこれがとても筋のよいアイディアであると思っている.DNAイベントレコーダーをつくるために必要な要素技術を考えるときにはビデオカメラのアナロジーがそのまま当てはまる.すなわちそれらは①DNAを情報記録媒体として利用する「メモリー(memory)」技術,②細胞内部 および外部状態を捉える「センサー(sensor)」技術,③センサーが捉えた状態情報をDNAメモリーに転写するゲノム編集などの「情報書込み(writer)」技術,④DNAに記録された情報から生命現象を再構築する「情報読出し(reader)」技術の4つである(図16-2).
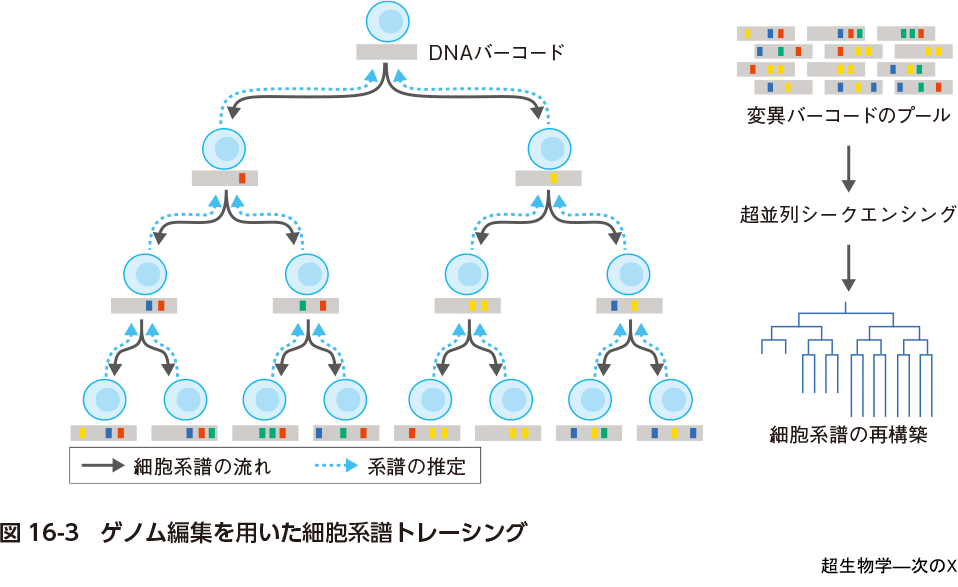
DNAイベントレコーディングの最もわかりやすい実装はゲノム編集(第1章)を用いた細胞系譜トレーシングである(第2章).これは今後すべてのDNAイベントレコーダーが備える基本機能になるであろう.細胞系譜トレーシングでは細胞内でゲノム編集(writer)によって標的とするDNAバーコード領域(memory)に経時的にランダムな変異が加えられ,蓄積していく変異は細胞分裂とともに娘細胞に引き継がれていく(図16-3).例えばこれが受精卵に仕掛けられた場合は発生後の組織の各娘細胞はさまざまな変異を標的DNA領域にもつことになるが,同じ母細胞から分裂した2つの娘細胞はきわめて似た変異パターンをもつことになる.したがって,組織からPCRによって標的DNA領域を増幅しこれをDNAシークエンシングで読み出せば進化系統樹推定と同じように観察時刻における標的DNA領域のパターンからそれらをもつ娘細胞の細胞系譜を予測できる(reader).このことは正に現存する生物種のDNA配列からそれらの進化系譜が推定できることと同じである.このような人工遺伝子回路はさまざまなシステムに実装することができ,ゼブラフィッシュ,マウスなどの脊椎動物の発生過程における細胞系譜を再構築しようとする研究2)〜4)や,がん細胞に仕掛けてその進展に寄与するゲノム変異と系譜を追う研究5)がある.さらに変異の入ったmemory DNA領域を転写させることによってscRNA-seqによって組織を構成する1細胞のトランスクリプトーム情報(セルタイプ)とその系譜を同時に読み出すこともできる(第3章,第4章).また空間トランスクリプトーム技術を用いれば,ある空間的な組織構造がどのように時間的に形成されていくか推定することもできるだろう(第14章,第15章).このとおり,細胞系譜トレーシングは特定の細胞状態を入力としてそのシグナルをwriterに渡すような明示的なsensorはもたないシステムであるが,それは確かに細胞系譜を感知して捉えており,DNAイベントレコーティングの好例である.
しかしながら,これまでの細胞系譜トレーシング技術には現在のままでは弱点がいくつもある.ひとつはwriterとして用いられるCRISPR–Cas9システムが染色体を切断するということである.細胞はDNA修復能をもつが完全ではない.染色体切断が細胞に負荷をかけることは疑いようがなく,これが分裂するすべての細胞にとめどなく起こるような状況で対象とする生命システムと細胞系譜トレーシング機構のどちらかあるいは両方が正しく作動するのかはよくわかっていない.加えて,Cas9タンパク質は標的DNAを切断後に頻繁に大きな欠失を引き起こすことが知られている.したがって,途中までうまく経時的に変異をmemory DNA領域に蓄積させられたとしても,突然そのmemoryが消失してしまうことも起こる.また特定のmemory領域にさまざまな変異を導入しようと複数箇所を一斉にターゲットすると同時に切断された箇所の間に挟まれる領域が丸ごと抜け落ちる.さらに,これまで発表された細胞系譜トレーシング手法で用いられたmemory DNAの情報許容量はとても小さく(1細胞あたり数百塩基対程度),理論的にも哺乳動物を構成する全細胞の系譜を1細胞分裂解像度で再構築できるようなものではない.実際にこれまでに同様の技術によって得られた細胞系譜情報はどれもきわめて粗い解像度のものである.最も重要なことに,再構成された細胞系譜が正しいものであるかという直接的な検証はこれまでにない.観察時刻における娘細胞が由来する組織情報やscRNA-seqによって同定されるセルタイプが細胞系譜と大きく矛盾しないという定性的な確認は行われているが,得られた細胞系譜の精度を定量的に評価した例はない.
このように細胞系譜トレーシング技術もまだ黎明期のものであるが,高度なDNAイベントレコーダーは上にあげた4つの要素技術をそれぞれどのように発展させていくとよいか考えると戦略が見えてくる.
❶ Memory
デジタル記録媒体と捉えることのできるDNAは画像や動画などの情報を保存でき,多くの人工DNAを用いると200メガバイトと大きな情報をとり扱うこともできる(第6章).細胞系譜トレーシングだけでなく,さまざまな高解像度のDNAイベントレコーディングを考えたときは,細胞の中にきわめて容量の大きなmemory DNAを準備することが必要である.しかしながら,長い人工的なDNAをmemoryとして染色体の特定領域(例えばマウスのROSA26)に導入するのは得策ではない.その理由は3つあげることができる.1つ目は,効率よく正確に長鎖DNAをシークエンシングする技術が実現する目処が立っていないことである.2つ目は,細胞特異的な染色体構造の特定の変化などによって,その領域の情報書込みや読出しが許されなくなってしまう可能性があることであり,3つ目は,先に述べたように,特定の領域に高負荷の書込みを行うとその領域自体が消失する可能性があることである.現在のインターネット上のクラウドサーバーシステムがそうであるように,リスクは多く分散させるのがよい.scRNA-seq技術と組合わせるとそのようなmemory DNAが実現しそうである.細胞内にある染色体のさまざまな箇所に小さなmemoryブロックが散っていて,これがRNAとして転写される状況を考える.このようにすると,細胞ごとに染色体上でバラバラの位置にある変異の組合せ情報はscRNA-seq技術とスケーラビリティーの高い短鎖DNAシークエンシング技術によって得ることができ,それぞれのmemoryブロックに独立に生じる多少の不具合(読み書きの阻害や消失)は深刻な問題にならない.十分に多いmemoryブロックは部分的な不具合を互いに補完できるだろうし,むしろ染色体の構造変化ヒストリーを捉えるようなものになるかもしれない.こう考えると問題は,そのようなmemoryブロック群はどのようなデザイン(人工DNA配列)であるべきか,どのように対象の機能を破綻させずに多くのmemoryブロックを染色体に導入した細胞あるいはマウスなどの個体を得るかという点になり,これが攻めるべき染色体工学,発生工学を中心とした研究課題ということになる.
❷ Sensor
すでに合成生物学は遺伝子発現制御,DNA組換え,タンパク質分解(第8章),RNA構造変化(第9章)によってさまざまな時間スケールの論理回路を実現しており,哺乳動物細胞におけるシグナル伝達を塩基編集として出力する系もある(第7章).一方で,自律的なそれらのDNAイベントレコーダーとしての実装はほとんど手が付けられていない.例えば,細胞が分裂とともに分化しどのようにそのセルタイプを変化させていくかは多くの生物学者の興味の対象である.scRNA-seqによるセルタイピングの成功が示すことは,任意の観察時刻において細胞それぞれのトランスクリプトーム状態(ヒトであれば約2万遺伝子の発現量)を知ることができれば,細胞の状態が定義できるということであった.また,2万遺伝子が成す2万次元ベクトル値がうまく次元圧縮できることは,実際には数十からせいぜい数百程度の遺伝子の発現状態情報(必ずしも高い定量性を必要としない)がセルタイプの規定には十分なことも明らかにした.ところが,このような複数の遺伝子の発現状態を同時にmemory DNAに書き込む決定的なアイディアはまだ生まれていない.Record-seqはタイプⅢ CRISPR familyのRT-Cas1–Cas2複合体を利用して大腸菌が自身のトランスクリプトームを逆転写してCRISPRアレイに取り込むことを集団のレベルで実現したが,単一の細胞レベルではその効率はきわめて低く,またRT-Cas1–Cas2あるいはCas1-Cas2複合体が哺乳動物細胞において外来DNAのCRISPRアレイへの取り込みを行えることを示した例はない(第7章).哺乳動物細胞では特定の遺伝子発現プロモーターによって塩基編集ツールを発現させ,特定のガイドRNA(guide RNA,gRNA)によってその遺伝子の発現状態をmemory DNA部分に記録する方法が提案されているが,この方法は複数の遺伝子に対する直行性をもたない.一方で,異なるgRNAを発現パターンの知りたい遺伝子のプロモーターでそれぞれ発現させるような方法は細胞内遺伝子発現プロファイルの経時的な記録を実現するかもしれない.また,任意のRNA配列をトリガーにできるような人工リボスイッチやMICRのようなマイクロRNAセンサー(第9章)は細胞状態の記録に利用できるかもしれない.このようなセンサーの信号情報が書き込まれたmemory DNA情報は細胞系譜情報をもつmemory DNAと組合わせると観察時刻に得た娘細胞群についてそれらの過去の遺伝子発現プロファイルの推定を可能にするはずである(ただし,ONスイッチだけでは2回目以降の同じ遺伝子発現イベントがある系譜以降のどこで生じたのかはわからないので,スパイクのような現象を捉えるためにはOFFスイッチと組み合わせて高感度に捉える必要がある).また遺伝子発現だけでなく,タンパク質間相互作用などさまざまな細胞内イベントをDNAの状態として出力するための技術基盤もある(第5章).
❸ Writer
今のところ情報書込みのためのwriterとしてはCRISPRゲノム編集系一択であろう.しかしながら,Cas9をこれに用いることの筋があまりよくないことはすでに述べた通りである.近年,他のイベントを記録する遺伝子回路では塩基編集ツール(BE:base editor)が用いられている例も出てきた6).BEはニッカーゼ型のnCas9あるいは不活性型のdCas9に脱アミノ化酵素を融合させたもので,Cas9と同様にgRNAによって標的配列に誘導することができる.BEにはシチジン脱アミノ化酵素によってgRNA標的配列のC→T編集を可能にするCBE(cytosine base editor)とA→G編集を可能にするABE(adenine base editor)がある.これらはDNA切断を介さずに標的配列に一塩基変異を導入できるために,一般にさまざまな細胞において毒性が低く,また野生型Cas9のように大規模な変異を起こしにくい(一定の頻度では欠失を起こす).しかしながら,誘導できる変異が不可逆であるために,例えばCBEを用いた場合はmemory DNA領域のシトシン(C)がすべてチミン(T)に変換され,塩基変換の飽和に至ると,情報記録能力も記録した情報も失ってしまうという欠点がある.これはmemoryブロック数が十分に多い細胞を準備すればよいとも考えられるが,実装への要求度が高くなる.これまでに進化生物学の分野ではDNAに可逆的な変異が何度も生じた配列からでも,うまく進化系統樹を再構築する手法が開発されてきた.私たちは2020年に入ってC→TおよびA→G編集活性を両方もつ新しいTarget-ACEmax(adenine cytosine editor)というBEを完成させた7).Target-ACEmaxは任意のアミノ酸をコードするコドンを別のものに変換できるBEの遺伝子治療分野などにおける可能性を拡張する.その一方で,私たちはこれが可逆的にC•G↔T•A塩基変換を誘導できるということがmemory DNAにおける塩基変換の飽和を緩和し,より拡張性の高いDNAイベントレコーディングを実現する可能性により大きな価値があると見ている.
❹ Reader
Memory DNAとしてブロック分散型のものを用いるとなると細胞ごとに染色体の異なる領域の情報を取得する必要がありscRNA-seqを用いるのがよいが,現在のscRNA-seq技術が取り扱える細胞の数は例えばマウスの成獣を構成する細胞をすべて取り扱えるほどにはなっていない.この点については楽観的になってゲノミクス分野における技術発展を待ちつつ,他のDNAイベントレコーダー関連技術の開発を進めても問題ないかもしれない.scRNA-seq技術のスケーラビリティーの急進は凄まじく,2015年にドロップレット型(第3章)の技術が数千〜数万細胞の解析を実現後,2020年にはスプリット・プール型の技術が400万細胞を取り扱えるようになったほどである8).ビデオカメラであればビデオデッキにあたるreader技術としてもう一つ重要なのは記録信号としてのDNA配列から生物学的イベントを再構成する計算機技術である.これについてもまだ多くの課題について手が付けられていない.例えば,これまでの細胞系譜トレーシング技術はそのほとんどが進化系統樹推定ソフトウェアを流用しているが,100万以上の入力配列からその系譜を推定できるソフトウェアはこれまでに開発されていない9).ヒトを構成する有核細胞数がざっくり見積もって5兆個あり,マウスでも数十億以上のオーダーであることを考えると,哺乳動物の全身構成過程をDNAイベントレコーダーで理解するためにはスーパーコンピューターとこれをうまく利用する系譜推定アルゴリズムの開発が必要である.このことは細胞系譜以外の細胞内イベントヒストリーを系譜情報とともに再構成する情報処理についても同様である.私たちはこの点についてもスーパーコンピューターを用いた新しい計算機技術フレームワークの研究を進めている.
最後に,ブロック分散型のmemory DNAをもつDNAイベントレコーダーではブートストラップ法によってreaderが再構成した情報の確からしさや,readerによる情報再構成精度の向上を狙うことができるという利点がある.これはmemoryブロック群が冗長に情報を記録できるように実装すると実現する.つまり,そのような実装ではある程度のイベントヒストリーは部分的なmemoryブロック群の標本からでも再構成できるはずであり,その標本とは全く独立の別のmemoryブロック群の標本からも情報を再構成すると,この2つの一致度を定量できる.あるいは,一致するようにイベントヒストリーの再構成手法を改良することができる10).例えば,完全に1細胞分裂解像度で細胞系譜を再構成できる十分な系においては,scRNA-seqについて娘細胞それぞれについて変異が導入されたmemoryブロック群すべての配列を得た後,2つに分けたmemoryブロック群それぞれから独立に得た2つの細胞系譜は完全に一致するはずである.
次のX
以上が私たちの「こんなことできたらいいな」であり,これが実現すれば生物学に大きなブレークスルーがもたらされることは想像できるかと思う.2000年代はじめにヒトゲノムが解読され,これが大きな知識の背骨として生物学の急進に貢献した.私たちはこれと同じように,一つの受精卵から全身がつくり上げられる様子を説明した(quick-and-dirtyなものではない)精密な地図を哺乳動物(マウス)について得られれば,これがまた次の生物学の背骨になると考える.もちろん,具体的にどうやるかという構想を無視して空想だけであれば,上にあげたDNAイベントレコーダーの夢はもっと広げることができる(ちょっとやってみよう).例えば発生の過程で細胞は互いにコミュニケーションしながら進展するので,異なる細胞の相互作用を捉え,その信号をDNAにレコーディングできるとよいが,どうやったらいいのかわからない.また,どれだけDNAイベントレコーディングが過去の状態を捉えられると言っても,アポトーシスなどによって死滅した細胞のことは捉えられない.進化系統樹を得るときと同じで,化石のように情報が凍結されたまま時空を飛び越えてこない限り,観察時刻まで脈々とmemory DNAが伝わってこないものについては「存在しない」ことと同等に扱われてしまう.細胞が死滅するときに隣の細胞にダイイングメッセージ(last signal)と自分のmemoryブロック群に刻まれた情報を受け渡すようにできればいいが,そんなことはどうやって実現したらいいのかさっぱりわからない(でも気づいてもらいたいのは,私が今ここで述べているDNAイベントレコーディングも10年前ならどうやって実現したらいいかだれも検討もつかなかったはずである).もしかするとCAR-T細胞に用いられるようなsynNotchなどはこれらの課題に使えるかもしれない(第10章).先人達がそうしてきたように,既にあるアイディアと技術や同じ時代を駆け抜ける研究者達が生み出すアイディアと技術を
また,上に述べたDNAイベントレコーダーであっても,つくってみないと見当もつかないことはたくさんある.例えば大腸菌では細胞のなかで人工的な遺伝子回路を作動させると小さなものであっても細胞分裂スピードが遅くなることが知られている11).これは大腸菌が本来発現させる必要のない遺伝子を発現させることで,ATPやリボソームなどのリソースが奪われるためである.大腸菌の場合は遺伝子発現の時間スケールと細胞分裂の時間スケールが近いためにこのような効果が見えるが,細胞周期の長い哺乳動物細胞で同様に外来の遺伝子導入によるリソース供給が問題になる例は知られていない(全身に蛍光を発現するマウスなどは誕生している).しかしながら,大規模なmemoryは追加で複製を必要とする染色体領域となるうえに,複雑なセンサーを細胞内に張り巡らせればそれを駆動させるだけのエネルギーは必要となるはずである.細胞系譜トレーシングだけのDNAイベントレコーダーであれば全身発生過程において問題なく作動するであろうが,現時点でどの程度の規模のものが生体システムの安定的なエネルギー供給を破綻させてしまうのか,あるいはそんなことは起こらないのか,わからない.いずれにしても,遺伝子工学や合成生物学では対象となる生命システムをうまく制御しようとしても一筋縄ではいかず,その過程で対象のことを深く知る機会に出会う.大規模なDNAイベントレコーダーを,例えば動物個体の規模で実装しようとする挑戦そのものが,私たちに細胞分化,発生や臓器と個体の機能について多くの知見を与えてくれるはずである.
参考文献
- Schiebinger G, et al:Cell, 176:1517, 2019
- Kalhor R, et al:Science, 361:doi:10.1126/science.aat9804, 2018
- Bowling S, et al:Cell, 181:1410-1422.e27, 2020
- McKenna A, et al:Science, 353:aaf7907, 2016
- Simeonov KP, et al:Cancer Cell:doi:10.1016/j.ccell.2021.05.005, 2021
- Anzalone AV, et al:Nat Biotechnol, 38:824-844, 2020
- Sakata RC, et al:Nat Biotechnol, 38:865-869, 2020
- Cao J, et al : Science, 370 : DOI:10.1126/science.aba7721, 2020
- Zou Q, et al:BMC Syst Biol, 11:100, 2017
- Masuyama N, et al:Curr Opin Chem Biol, 52:63-71, 2019
- Scott M, et al:Science, 330:1099-1102, 2010