
レビュー編 Ⅱ.基礎知識
3 Low-complexity配列による相分離・相転移機構
加藤昌人
(テキサス大学サウスウエスタンメディカルセンター生化学科/量子科学技術研究開発機構量子生命科学研究所)
はじめに
真核細胞内には,ミトコンドリア,小胞体などの脂質二重膜をもっているオルガネラ以外に,膜をもたない構造体(非膜性オルガネラ)も多数存在している.その代表例は,細胞質に形成されるRNA顆粒である.RNA顆粒は,RNAと多くのRNA結合タンパク質のダイナミックな凝集体であり,膜がないため短時間に出現,消失したり,顆粒同士で融合したりできる.RNA顆粒が,外界と仕切る膜もなしに,物理化学的にどのように形成維持されているのかは,古くからの生物学の謎であった.しかし,これまでRNA顆粒に局在すると確認されたRNA結合タンパク質の多くにはLCD(後述)が含まれており1),LCDがRNA顆粒の形成機構に重要な役割を果たしていることがわかってきた.
通常,構造をもつタンパク質は20種類あるアミノ酸すべてを駆使した複雑な配列をもつ.しかし,遺伝子解析の技術発展に伴ってタンパク質配列の解析が容易になってくると,限られた種類のアミノ酸ばかりが頻出する奇妙な配列領域をもつタンパク質が多数見つかってきた.そのような配列領域はLCDとよばれ,配列の特性上安定した構造をもたない天然変性領域としても認識されている.50残基以上の長さのLCDは,ヒトのプロテオームの10%以上のタンパク質に含まれると予想されている2).それらの多くは,細胞内で重要な機能をもつ制御タンパク質である.前述したRNA結合タンパク質ばかりでなく,多くの転写因子の活性に必須な活性化ドメインもLCDである.例えば,酵母Gal4転写因子とヘルペスウイルスのVp16転写因子の活性化ドメインは,酸性アミノ酸(アスパラギン酸とグルタミン酸)がくり返し出現するLC配列である3)4).他の例では,ヒトのRNAポリメラーゼⅡのC末端には,7残基配列(YSPTSPS)が52回もくり返されるLCDがあり,このくり返し配列内のセリンとスレオニンのリン酸化を通して遺伝子の転写開始が制御されている5)6).また,mRNA成熟機構で働く多くのスプライシング因子は,アルギニン(R)−セリン(S)リピートドメイン(RSドメイン)をもち,これらのドメインがスプライシング活性に必要不可欠のLCDであることがわかっている7).
このように,LCDには重要な機能が備わっていることがわかっている.しかし,構造をもたないLCDは分解されやすく,凝集・沈殿しやすいため,生化学的・構造生物学的な実験には扱いにくく,これまで研究対象としては敬遠され,その機能解析はほとんど進んでいなかった.しかし,近年,FUSやhnRNPA2などのRNA顆粒に局在するRNA結合タンパク質のLCDが,凝集してRNA顆粒を模倣するハイドロゲルを形成することが報告された1).これを契機に,じつは凝集(自己相互作用)しやすい性質それ自体が,LCDの細胞内での本来の機能であることがわかってきた.LCDの相分離・相転移は,その機能を利用した代表例であり,RNA顆粒などの細胞内非膜性構造体の形成原理と考えられている.また,TDP-43やFUSのLCDには,家族性の筋萎縮性側索硬化症(ALS)を引き起こす変異が多数同定されている8).多くの変異は,LCDの自己相互作用を必要以上に促進し,細胞内にこれらのタンパク質の線維凝集体を形成・蓄積させることが病原機構であると考えられている9).
ここでは,LCDの自己相互作用によって誘導される相分離・相転移がどのように起こるのか,またどのように制御されているのかを,次章以降のプロトコールを理解する助けとなるようまとめておきたい.
LCDの相転移―ハイドロゲル形成
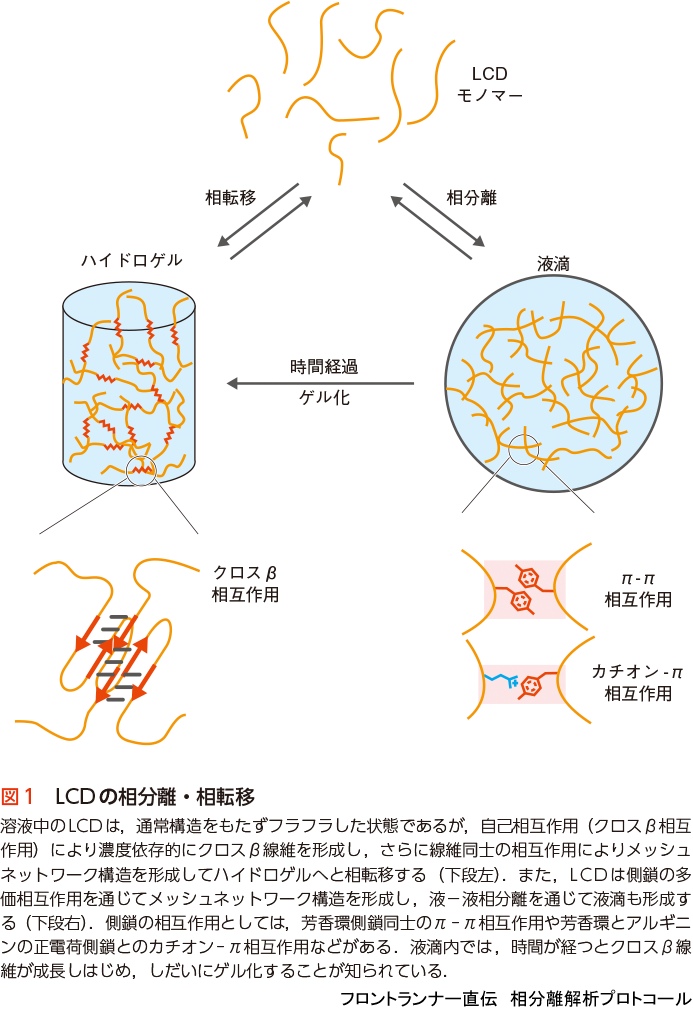
細胞内の非膜性構造体であるRNA顆粒には,多くのRNA結合タンパク質が蓄積する.じつは,それらのRNA結合タンパク質の大半がLCDをもっていることがわかっている1).RNA顆粒に蓄積するFUSやhnRNPA2のLCDは,濃度依存的に溶液状態からハイドロゲル(hydrogel)状態に“相転移”することが示された(図1)1)10).この現象を利用したハイドロゲル結合アッセイ法(プロトコール編6参照)を用いて,これらのハイドロゲルが,RNA顆粒のように他のタンパク質のLCDを蓄積することが示された1).電子顕微鏡で観察してみると,ハイドロゲルは無数のアミロイド様線維(クロスβ線維)がより集まってできていることがわかった(図1)1).線維が伸長するにつれ,線維同士が接触して絡み合うことで,線維の立体的メッシュネットワーク構造が形成され,溶液からゲル状態に相転移したのである.アミロイド線維は,神経変性疾患などの病原性線維凝集体として知られており,その特徴は,アミロイド線維共通のクロスβ構造をもち,かつ事実上不可逆的とも言える高い安定性である11).しかし,FUSやhnRNPA2のLCDのアミロイド様線維は,同様なクロスβ構造をもつが,40˚C程度に加熱したり,低濃度のSDSで処理するだけで解離するほど不安定である1)12).固体NMR構造解析から,FUS LCD線維のクロスβコアには疎水性残基がほとんどないことがわかり,一方,病原性線維のコアでは多くの疎水性残基が相互作用してコア構造を安定化しており,この違いがLCDのクロスβ線維の不安定さの理由であることが明らかになっている13).つまり,LCDのクロスβ線維形成は可逆的であり,細胞内で機能するダイナミックなRNA顆粒の構成要素として考えるのに問題ないことがわかった.
細胞内に存在するLCDのクロスβ線維
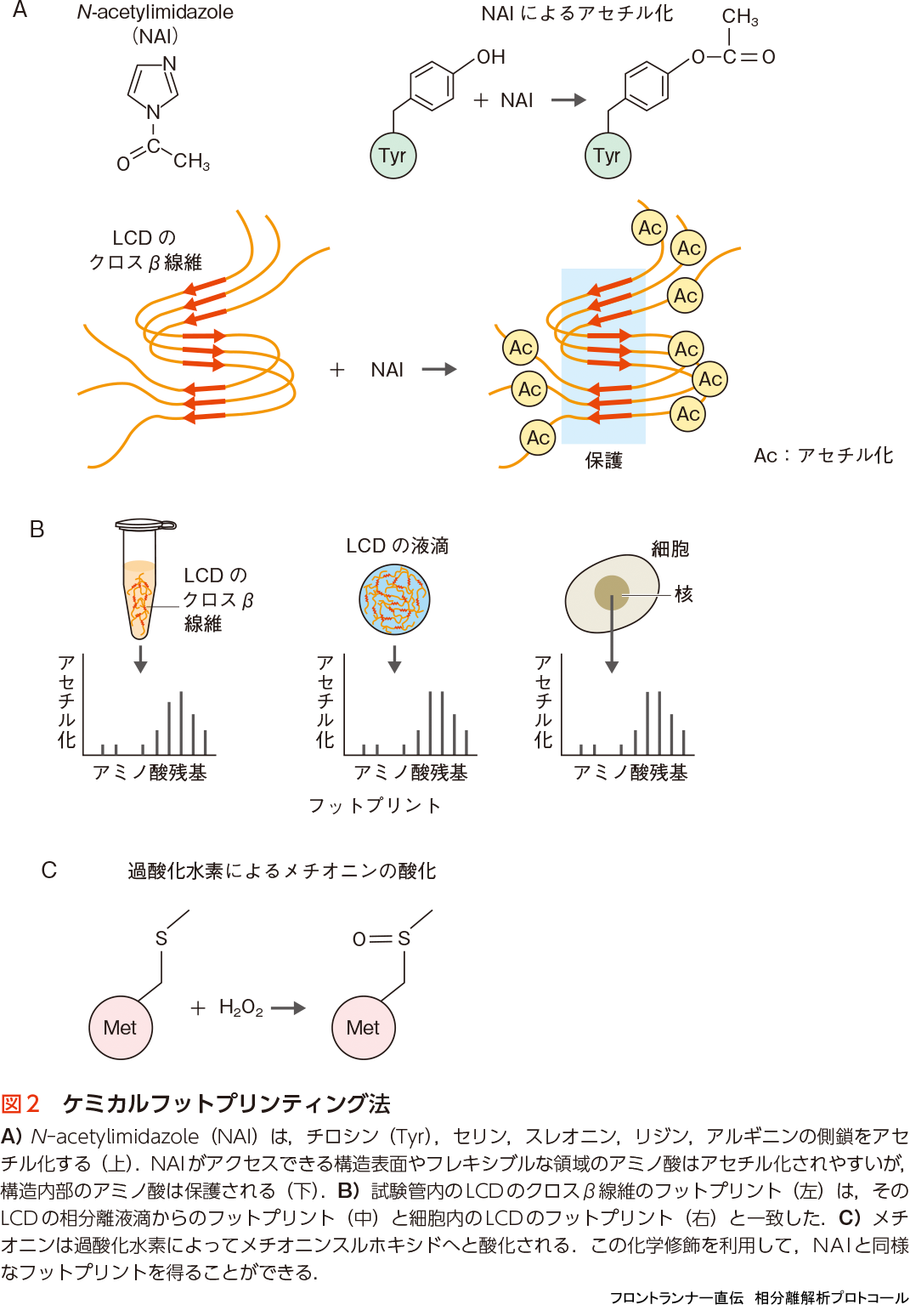
LCDのクロスβ線維が細胞内に存在している証拠は,ケミカルフットプリンティング法により示されている.タンパク質を特定の低分子化合物で化学修飾すると,立体構造内のアミノ酸残基は化学修飾から保護されるが,構造表面の溶媒に露出したアミノ酸残基はよく修飾を受ける.質量分析によりアミノ酸配列上の保護部分と修飾部分のパターンを解析したものを,“フットプリント”とよぶ.例えば,低分子化合物のN-acetylimidazole(NAI)は,特定のアミノ酸の側鎖を生理条件下でアセチル化することができる(図2A).このNAIのアセチル化によるhnRNPA2 LCDの試験管内のクロスβ線維の“フットプリント”と,細胞の核内に存在するhnRNPA2のLCDの“フットプリント”がよく一致することが示された(図2B)14).また,メチオニンの過酸化水素水(H2O2)による酸化を利用した方法もある.メチオニンは20種のアミノ酸のなかでも最も酸化されやすく,酸化によってメチオニンスルホキシドになる(図2C).10個のメチオニンをもつTDP-43 LCDのクロスβ線維を安定同位体過酸化水素(H218O2)で酸化し,質量分析によって各メチオニンの酸化パターンを解析することによって,NAIと同様に,クロスβ線維の“フットプリント”が得られる.そのフットプリントが,細胞内のTDP-43から得られたフットプリントと一致することが示された15).これらの結果は,細胞内のLCDも,試験管内のLCDのクロスβ線維と同じ構造をもっていること,すなわち細胞内にもLCDのクロスβ線維が存在していることを示唆している.
LCDの液−液相分離―液滴形成
FUSやhnRNPA1/A2などのLCDは,液−液相分離を通じて,ダイナミックな液滴(liquid-like droplet)を形成することもできる(図1)16)〜20).この液滴は,外部の力で形を変えたり,液滴同士で融合したり,他のLCDやDNA/RNAを取り込んで蓄積することができ18)19),細胞内の非膜性構造体と同様なふるまいをすることから,液−液相分離が細胞内非膜性構造体の形成機構として考えられるようになった.相分離液滴内では,構造をもたずフレキシブルなLCDをもつ分子同士が,アミノ酸側鎖の弱い結合力による多価(1分子が多数の他分子と同時に)相互作用をして,三次元のメッシュネットワークを構築していると考えられている(後述,図1).これらの弱い結合は,くっついたり離れたりをくり返しているため,分子は液滴内を動き回ったり,液滴の内外を行き来したりしている.
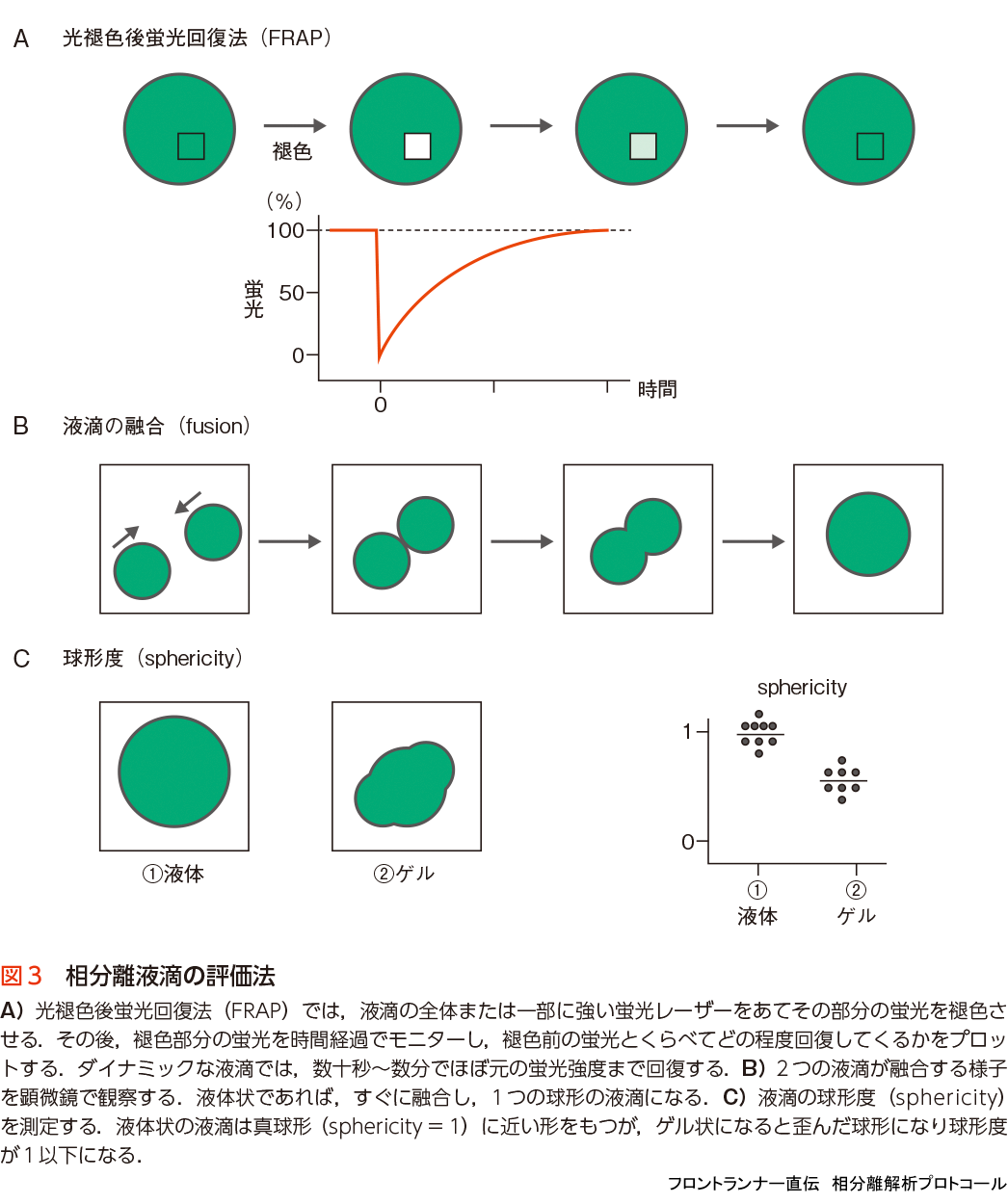
液滴が実際にどれだけ液体状であるかを測定するために,光褪色後蛍光回復法(fluorescence recovery after photobleaching:FRAP)がよく使われる.これは,蛍光ラベルした液滴の一部または全体をレーザー光で蛍光褪色させた後,褪色領域の蛍光が回復してくる時間を測定する方法で,液滴は短時間(数十秒〜数分)でほぼ元の蛍光強度まで回復することが示されている(図3A)18).この方法は,細胞内の非膜性構造体が液体状かどうかを調べるのにも有効である20).その他の指標としては,2つの液滴が融合する様子を観測する方法がある.液体状の液滴は,別の液滴と接触するとすばやく融合し1つの液滴となる(図3B)18).また,融合後の液滴の球形度(sphericity)も指標として使われており,真球に近い形に戻るほど液体状であることを示す(図3C)18).
相分離のためのLCDのアミノ酸配列特性
構造をもつタンパク質は,変性して構造を失うとほとんどの場合アモルファス(無定形)凝集体を形成して沈殿してしまう.もともと構造をもたないLCDも,同様の傾向をもつ.しかし条件しだいでは,LCDはアモルファス凝集体ではなく,前述したように相分離・相転移を通してダイナミックな液滴や可逆性のクロスβ線維を形成することができる.では,どのようなアミノ酸配列の特性があればアモルファス凝集せずに相分離を起こすのだろうか? 以降,最もよく研究されているFUSとhnRNPA1のLCDを代表例として話を進める.
これまでの研究から,LCDによる相分離機構の本質は,LCDが“多価相互作用(multivalent interaction)”することによって,“三次元メッシュネットワーク構造(3D mesh network structure)”を形成することであることがわかってきた(図1).ここで言う多価相互作用とは,1つのタンパク質分子が他の複数の分子と同時に相互作用することである.メッシュネットワーク構造を形成するためには,原理的には3カ所(3価)以上の結合点を1分子内にもつことが必要である(図4A)21).つまり,1つの分子が,異なる3分子(かそれ以上)と同時に結合し,その多価相互作用がほとんどの分子で同時に起これば,おのずとメッシュネットワーク構造が構築されることになる.
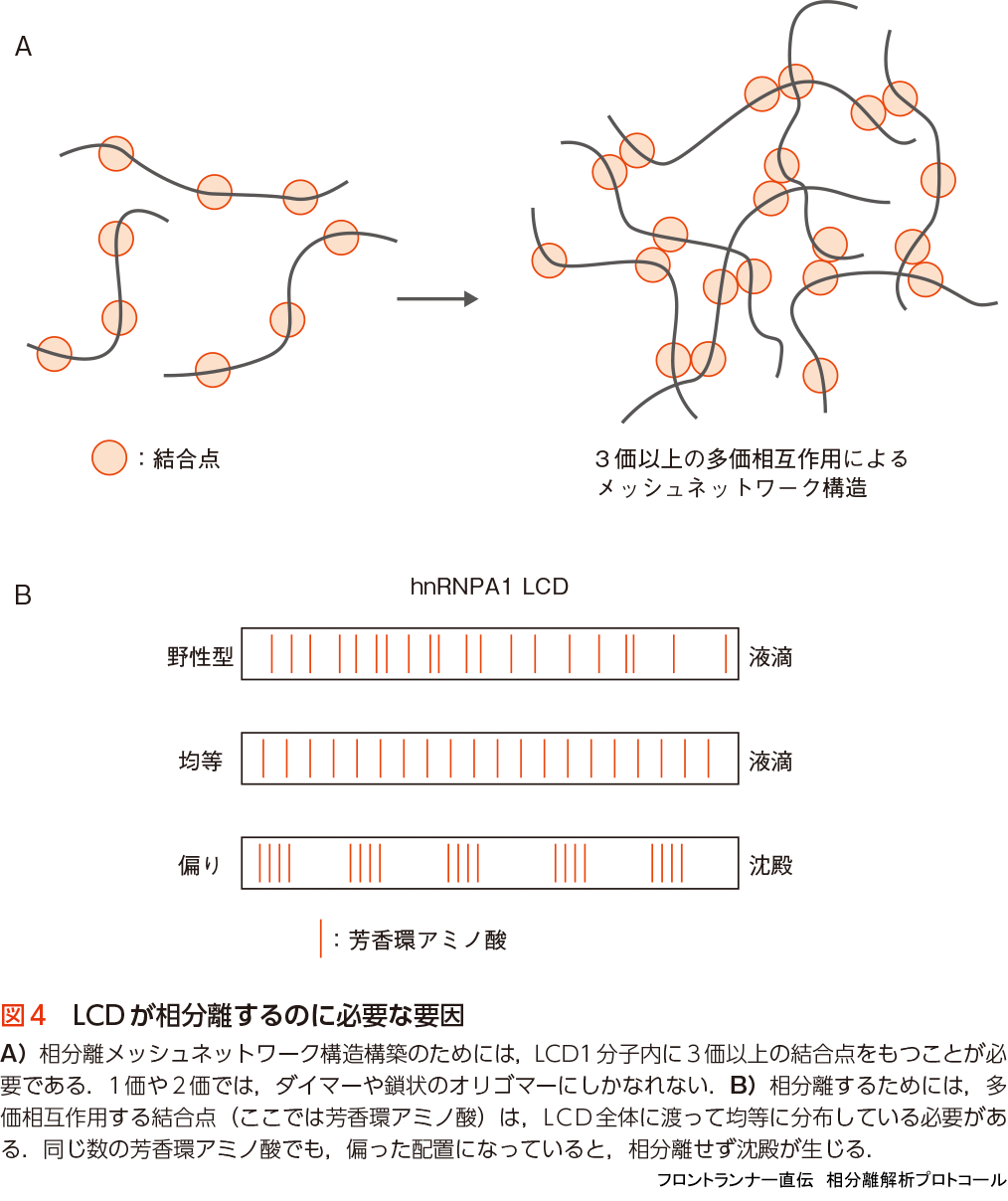
多価相互作用するための結合点には,主にアミノ酸側鎖の弱い結合が使われる.側鎖の弱い結合とは,チロシンやフェニルアラニンの芳香環側鎖同士の相互作用(π - π interaction)や,同じく芳香環側鎖とアルギニン側鎖の正電荷間で起こる相互作用(cation- π interaction,図1),正電荷側鎖と負電荷側鎖間の静電相互作用などである.FUSとhnRNPA1のLCDには,チロシンとフェニルアラニンがくり返し出現し(FUS 27個,hnRNPA1 20個),これらのアミノ酸の芳香環側鎖同士の相互作用(π - π interaction)が結合点として働くことが示されている.1アミノ酸だけの相互作用は弱く,それだけではすぐに離れてしまうが,これだけの数の結合点が1分子内にあれば,そのうちの複数の結合点がいつも他の分子と相互作用していて,分子は液滴内で動きまわりつつも,メッシュネットワーク構造内にトラップされ続けることになる.
また,結合点としての芳香環アミノ酸の数(価数)だけでなく,それらの配列上の位置も相分離に重要な要因である.FUSの27個のチロシンは,アミノ酸配列上であまり密集せず,比較的均等に配置されている.hnRNPA1においても同様な傾向がみられる(図4B).実際,芳香環アミノ酸を完璧に均等配置させたhnRNPA1のLCD変異体は,野生型と変わらず相分離液滴を形成するが,芳香環アミノ酸の数はそのままで偏った配置にした変異体LCDは,相分離せずにアモルファス沈殿になった(図4B)22).このように,結合点が互いに密集せず比較的均等に配置されていることが,相分離にとって重要であることがわかっている.
LCD以外の相分離ファクター
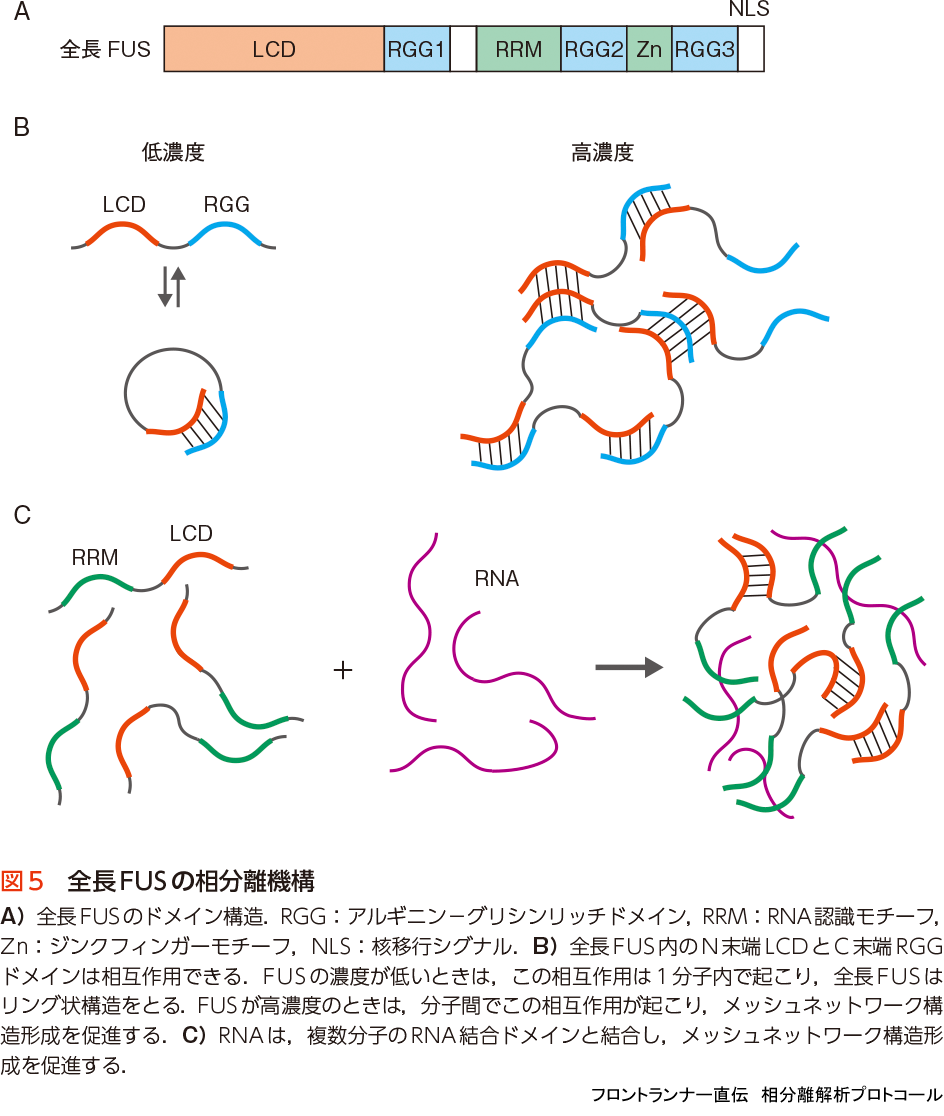
全長のFUSは,N末端のLCD以外にも,2つのRNA結合ドメインと3つのRGGドメイン(アルギニンとグリシンに富むLCD)をもつ(図5A).全長FUSも相分離することができ,それにもN末端のLCDが重要な駆動力であることに変わりはないが,以下に解説するように,RNA結合ドメインとRGGドメインも重要な働きをすることがわかっている.
前述したようにアルギニンは,チロシン側鎖の芳香環とカチオン- π 相互作用する.そのため,C末端側のRGGドメインは,チロシンに富むN末端のLCDと相互作用することができる.FUSの濃度が低い溶液中では,LCD-RGG間の相互作用は一分子内だけで起こり(intramolecularly),FUSはリング状構造をとることが知られている23).しかし,タンパク質濃度が高くなると,この相互作用が分子間で起こるようになり(intermolecularly),LCD-LCD間の相互作用に加え,LCD-RGG間の相互作用が相分離のためのメッシュネットワーク構造構築に寄与する(図5B).そのため,全長FUSが相分離するのに必要なタンパク質濃度は,LCDのそれよりもかなり低く,FUSの細胞内での濃度(2〜8μM)とほぼ同程度である18).
RNA結合ドメインもRNAの存在下で,メッシュネットワーク構築のための多価相互作用を促進する.これはRNA結合ドメインを通して結合したRNAに,別の分子のRNA結合ドメインが結合することができるので,タンパク質間をさらに架橋することによってメッシュネットワーク構造構築を促進するからと考えられる(図5C).ところが,RNAの量がタンパク質の量よりも過剰になると,一転して相分離は阻害されてしまう24).これは,タンパク質が過剰な量のRNAに分散して結合するため,タンパク質同士の相互作用が阻害されるためだと考えられる.
クロスβ相互作用と液滴のハイドロゲル化
先に述べたように,FUSやhnRNPA1のLCDは,濃度依存的にクロスβ(アミロイド様)線維を形成することができる1)25).線維内では,各分子はアミロイド様線維特有のクロスβ構造をとり,隣接サブユニット同士で相互作用している.これはとても特異的な自己相互作用で,クロスβ相互作用とよぶ.相分離液滴内で,数分子のLCDでもクロスβ相互作用で短い線維を形成すれば,それはメッシュネットワーク構造形成上の分岐点となり,相分離を促進させる(図1).実際,液滴内でのLCDのクロスβ構造の存在は,前述のケミカルフットプリンティング法によって示されている(図2)14)15).
FUS LCDやhnRNPA1 LCDの相分離液滴は,形成後しばらくはダイナミックな液体状態を示しているが,古くなった液滴では,蛍光褪色後の回復が遅くなったり,元の蛍光強度に戻らなくなったりする.また,液滴同士が融合しても真球形に戻らなかったり,接触しても融合しなくなることさえある18).これらの変化は,液滴内部で形成されはじめたLCDの短いクロスβ線維が時間とともに成長し,線維同士の相互作用が起こりはじめ,液体状態からハイドロゲル状態へと変化した結果と考えられている(図1).実際,さらに放置しておくと,液滴から放射状に線維が成長してくるのが観測されている18).
クロスβ線維の形成は濃度依存的であるので,タンパク質が凝集されて濃度が高くなっている液滴内部は,もともと線維形成が促進される環境にある.細胞内では,それを制御するメカニズム(翻訳後修飾やシャペロン)が存在しているが(後述),試験管内の液滴ではそのメカニズムがないため,時間とともにクロスβ線維が伸長し,ゲル化へと至ると考えられている.つまり,細胞内でもその制御システムに異常をきたしたり,LCDの変異によって線維形成が必要以上に促進されたりすることによって,非膜性構造体をもとにして線維凝集体が形成されることが,ALSなどの神経変性疾患の病原メカニズムの1つではないかと推察されるようになっている26).
細胞内相分離の制御機構
細胞内には,非膜性構造体を短時間に形成・消滅させるために,また,クロスβ線維の異常成長を防ぐために,LCDの自己相互作用の制御機構が存在する.その1つは,翻訳後修飾(post-translational modification:PTM)である.構造をもたないLCDは,構造をもったタンパク質よりもPTMを受けやすい27).LCD上の特定のアミノ酸のPTMによる化学状態の変化は,その自己相互作用に影響を与えるため,PTMは重要な相分離の制御機構として認識されている.
例えば,リン酸化されたアミノ酸はリン酸基の2価の負電荷の影響で溶解度が上がる.また,リン酸基のかさ高さもアミノ酸同士の相互作用を阻害する.それゆえFUS LCDのリン酸化はクロスβ相互作用を阻害すると同時に,液滴形成も阻害することが示されている13)28).一方,正電荷を帯びたアルギニンの側鎖がメチル化されると,その正電荷は下がる.その電荷の違いによって,前述したような全長FUSのRGGドメインのアルギニンとLCDのチロシンとのカチオン-π相互作用の強弱が変化し,FUSの相分離が制御されていることもわかっている29).さらに,メチオニンは,ミトコンドリア活性の副産物である活性酸素種(H2O2等)によって,溶解度の高いメチオニンスルホキシドに酸化される.一方,細胞内にはメチオニンスルホキシドを特異的に還元してメチオニンに戻すメチオニンスルホキシド還元酵素が存在する.つまり,細胞内にはメチオニンの酸化・還元サイクルによって,タンパク質の活性を制御するシステムが存在しているのである.酵母のAtaxin-2やTDP-43のLCD内のメチオニンは,他のタンパク質でのメチオニンの平均的な頻度よりも高い頻度で出現しており,そのメチオニンの酸化・還元状態によって,これらのLCDの液滴形成(還元)と溶解(酸化)が制御されていることが明らかにされている15)30). LCDの自己相互作用は,シャペロンによっても制御されている.ヒートショックタンパク質のHsp27は,FUSのLCD全体に渡って弱く結合してFUSの相分離を阻害することが示された31).同じくヒートショックタンパク質のHSPB1も,TDP-43のLCDに直接結合し,液滴内でのTDP-43 LCDのクロスβ線維形成を阻害することが示された32).さらには,インポーティンβファミリーのKapβ2は,FUSのC末端にある核移行シグナルペプチドに結合し,核膜孔を通じてFUSを核内に輸送する役割をもつが,この結合もシャペロン作用として働き,FUSの相分離を阻害することが報告された33).
おわりに
ここでは,LCDの自己相互作用による相分離・相転移に着目して議論してきた.これまでのところ,この自己相互作用の駆動力として議論されているのは,アミノ酸側鎖の弱い相互作用ばかりで,ポリペプチド鎖の主鎖による相互作用(ペプチド結合間の水素結合)については議論されていない.タンパク質1分子がフォールディングして構造を形成するとき,側鎖だけの相互作用だけで最終的な構造に到達するわけではない.タンパク質がアミノ酸の側鎖と主鎖の両方をもつ以上,タンパク質分子の会合,特にLCDが多価相互作用して絡み合うときには,主鎖と側鎖の両方の相互作用の影響を考慮しなければならないと考えられる.実際,最新のわれわれの研究からは,TDP-43 LCDの主鎖間の水素結合1つの増減により,相分離状態が変化することが示されている34).LCDの機能解析の研究はまだ途についたところである.今後主鎖の相互作用も考慮に入れた研究も行われ,LCDの相分離・相転移機構のさらなる解明が進むことを期待する.
文献
- Kato M, et al:Cell, 149:753-767, 2012
- van der Lee R, et al:Chem Rev, 114:6589-6631, 2014
- Ma J & Ptashne M:Cell, 51:113-119, 1987
- Triezenberg SJ, et al:Genes Dev, 2:718-729, 1988
- Corden JL, et al:Proc Natl Acad Sci U S A, 82:7934-7938, 1985
- Egloff S, et al:Trends Genet, 28:333-341, 2012
- Wang J, et al:Genes Dev, 10:2588-2599, 1996
- Bentmann E, et al:FEBS J, 280:4348-4370, 2013
- Mackenzie IR, et al:Lancet Neurol, 9:995-1007, 2010
- Han TW, et al:Cell, 149:768-779, 2012
- Makin OS, et al:Proc Natl Acad Sci U S A, 102:315-320, 2005
- Kato M & McKnight SL:Proc Natl Acad Sci U S A, 118:doi:10.1073/pnas.2114412118, 2021
- Murray DT, et al:Cell, 171:615-627.e16, 2017
- Xiang S, et al:Cell, 163:829-839, 2015
- Lin Y, et al:Proc Natl Acad Sci U S A, 117:28727-28734, 2020
- Elbaum-Garfinkle S, et al:Proc Natl Acad Sci U S A, 112:7189-7194, 2015
- Nott TJ, et al:Mol Cell, 57:936-947, 2015
- Patel A, et al:Cell, 162:1066-1077, 2015
- Lin Y, et al:Mol Cell, 60:208-219, 2015
- Molliex A, et al:Cell, 163:123-133, 2015
- Martin EW & Holehouse AS:Emerg Top Life Sci, 4:307-329, 2020
- Martin EW, et al:Science, 367:694-699, 2020
- Hamad N, et al:Chem Commun (Camb), 56:9134-9137, 2020
- Maharana S, et al:Science, 360:918-921, 2018
- Kato M, et al:Methods, 126:3-11, 2017
- Pakravan D, et al:J Mol Cell Biol, 13:15-28, 2021
- Xie H, et al:J Proteome Res, 6:1917-1932, 2007
- Monahan Z, et al:EMBO J, 36:2951-2967, 2017
- Qamar S, et al:Cell, 173:720-734.e15, 2018
- Kato M, et al:Cell, 177:711-721.e8, 2019
- Liu Z, et al:Nat Struct Mol Biol, 27:363-372, 2020
- Lu S et al:bioRxiv, doi:10.1101/2021.10.14.464447, 2021
- Yoshizawa T, et al:Cell, 173:693-705.e22, 2018
- Zhou X, et al:Science, 377:doi:10.1126/science.abn5582, 2022