
概論
マクロファージ学〜その歴史と現在地
Studies of macrophage
佐藤 荘
Takashi Satoh:Laboratory of Host Defense, World Premier Institute Immunology Frontier Research Center, Osaka University/Department of Host Defense, Research Institute for Microbial Diseases (RIMD), Osaka University(大阪大学免疫学フロンティア研究センター自然免疫学/大阪大学微生物病研究所自然免疫学)
最近,免疫学の大きなトピックの1つとしてマクロファージが挙げられる.phagocyteは無脊椎動物において異物を貪食する細胞としてMetchnikoffによって発見され,そのなかから単核のものを“マクロファージ”と名付けた.当初は,体内に侵入した異物やごみを処理するしか仕事をしない細胞だと思われており,免疫学でもスポットライトの当たらない補欠の細胞であった.しかし,近年のM1・M2のコンセプトから,最近のマクロファージサブタイプ(亜種)の研究が徐々に増えつつあり,2011年からはPubMedでの“macrophage”でヒットする論文は毎年10,000を超え続けている.長年スポットライトの当たっていなかったマクロファージ研究は,今まさにNEON GENESIS(新世紀)を迎え,その“序”の章がスタートした.本稿では,前半にてマクロファージの分化や種類の多様性について,そして後半では最近のさまざまなマクロファージ研究のテーマのなかでも,“疾患”との関係にフォーカスを当て概説する.
1マクロファージの起源と分化
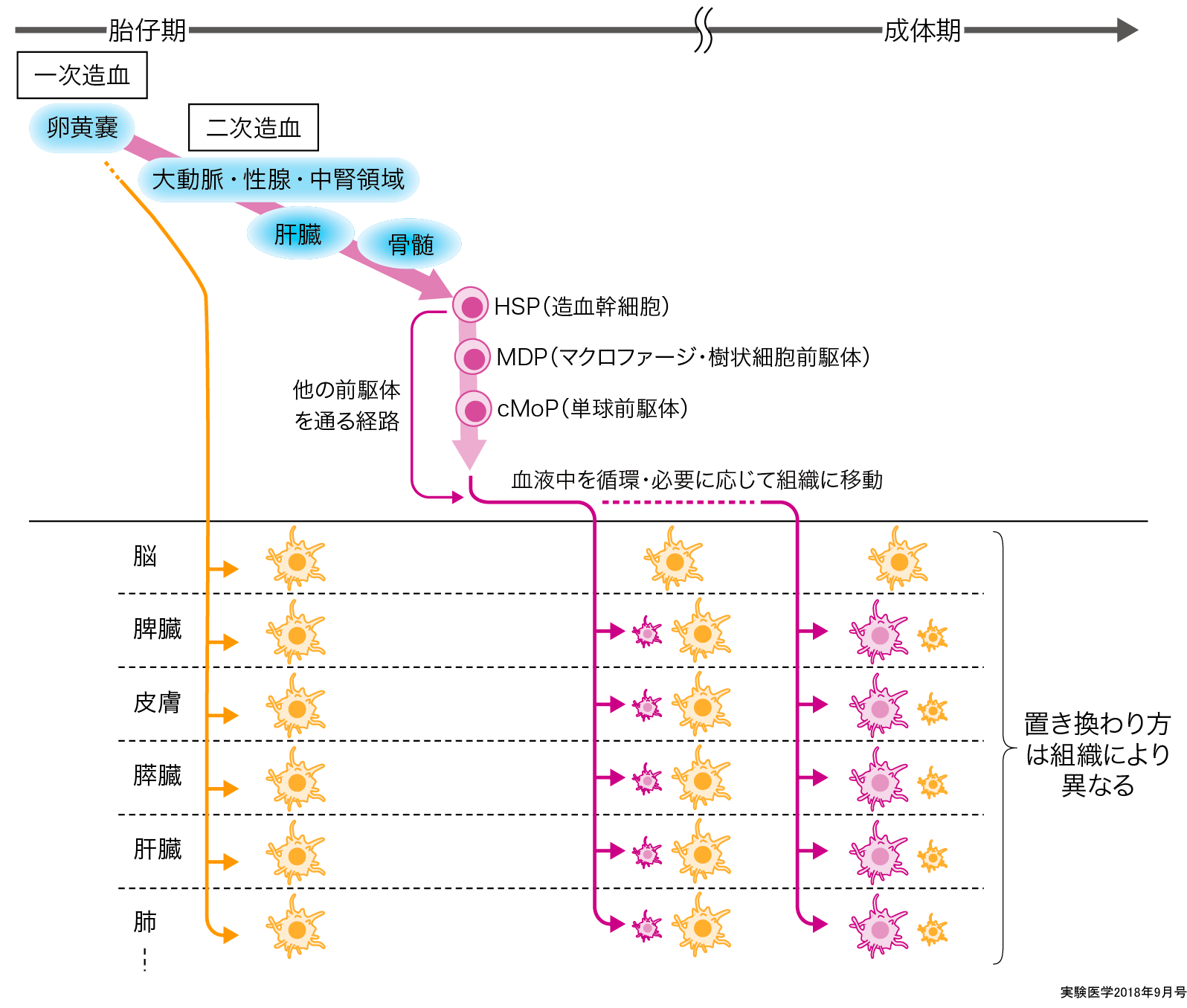
現在,マクロファージの起源は胎生期の卵黄嚢由来で,生体になった後も組織に分布しているマクロファージと,骨髄の前駆体由来でそれが組織に行って常在型となるマクロファージとに分けられる.この歴史について繙いていく(図1).
1968年にFurthやCohnらを中心とした研究者らは,前駆体から単球が出現して血中に流れ,その血中に漂っている単球が必要に応じて末梢組織に移動し,マクロファージに分化するというmononuclear phagocyte system(MSP)という概念を提唱した2)3).一方で,造血系は卵黄嚢の一次造血から大動脈・性腺・中腎領域での二次造血が行われ,そこではじめて血管内皮細胞から造血幹細胞が発生し,そして胎仔の肝臓から骨髄へと造血の場が移行していく.リンパ球など他の免疫細胞と同様に単球はこの肝臓や骨髄の造血幹細胞から発生するが,単球から分化するはずのマクロファージは,その単球が造血幹細胞から発生するよりも前の段階の内胚葉由来卵黄嚢の血島に出現することから,このMSPという概念は一部破綻が生じているとも考えられる.近年,
また,マクロファージの前駆体からの分化に関しても近年研究が急速に進んでいる.マクロファージおよび樹状細胞はHSPの下流にあるmacrophage DC progenitor(MDP)7)から派生することをGeissmannらが報告し,さらにFeuererらのグループはその下流に単球の前駆体であるcommon monocyte progenitor(cMoP)があることを証明した8).最近,樗木らのグループはヒト臍帯血や骨髄中にこのcMoPのヒトカウンターパートが存在していることを明らかにし9),マウスで展開されてきていたマクロファージの前駆体研究がヒトでも示された大きな一歩である.一方でMDP以外にもマクロファージ・単球の前駆体が存在していることも証明されている10).分化や活性化についても,マクロファージの種類によって未解明な点は多い.
2M1/M2という概念
発見から1世紀近くもの間,マクロファージは1種類しかないと考えられてきた.最近よく聞くM1/M2マクロファージというものも,状態を示したものでマクロファージの種類ではない.ここではM1/M2の概念について紹介したい.
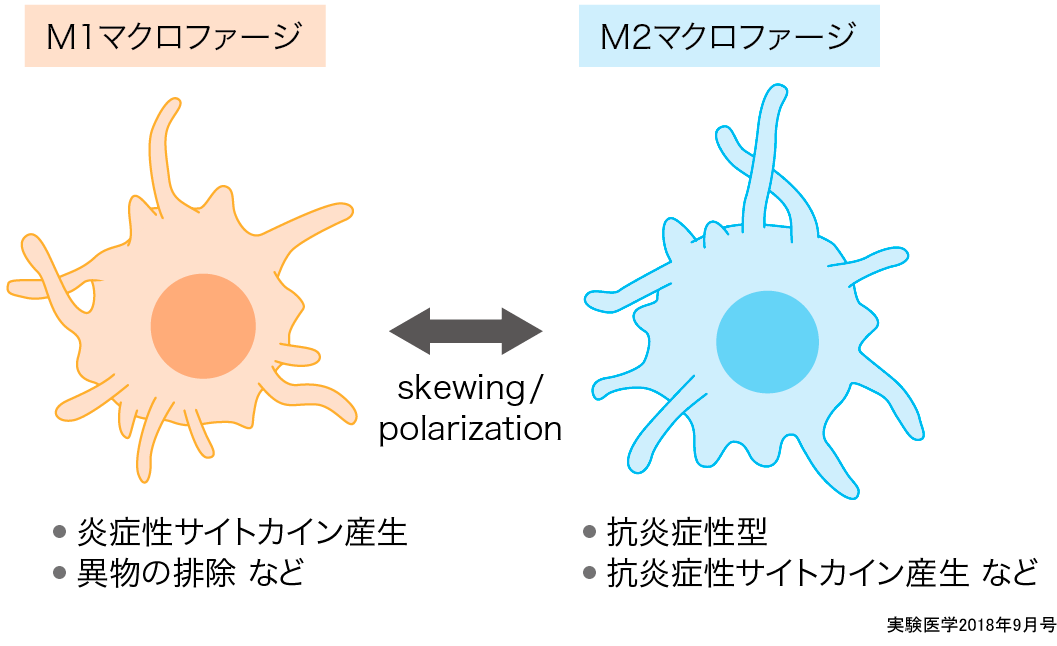
1990年代にGordonらは,それまでに報告されていた病原体に対するマクロファージの活性化とは異なる活性化(alternative activation)がIL-4によって起きると最初に報告し11),その後2000年頃にHillらがM1,M2マクロファージの考え方を提唱した(図2)12).それに続きGordonやSicaらはM1マクロファージはIFN-γとLPSなどの菌体成分などによって活性化(classically activation)したマクロファージで炎症性サイトカインを強く発現し,バクテリアを殺傷する役割を果たしている一方で,M2マクロファージはIL-4もしくはIL-13によって誘導され,抗炎症作用をもつという,マクロファージは1種類ではあるが,2つの状態を行き来する(polarization・skewing)という考えを提唱した13).最近のマクロファージ研究のなかではこのM1,M2という言葉がよく使われているが,これらはもともと
またこのM1,M2の
3さまざまな疾患にかかわるマクロファージのサブタイプ
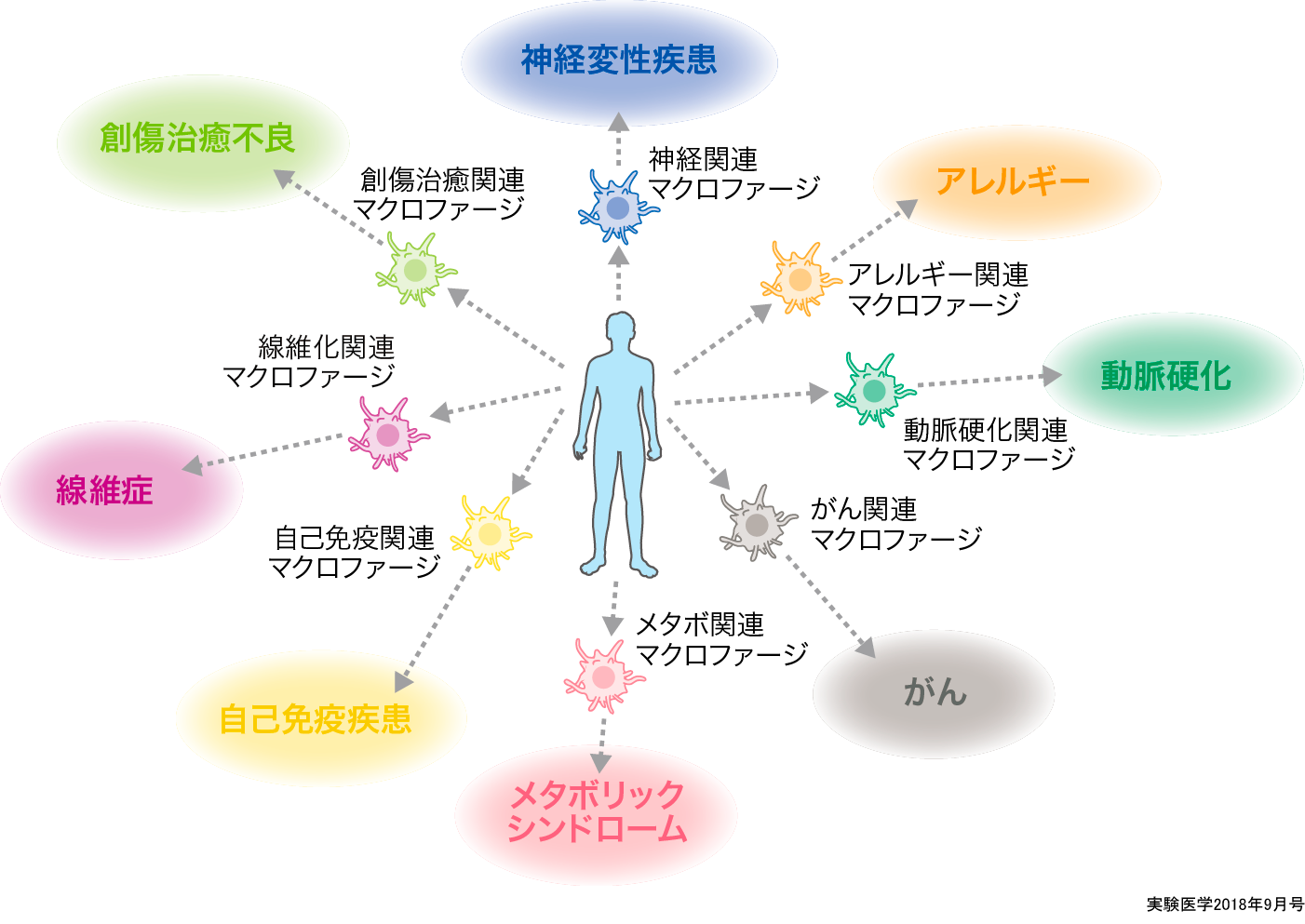
マクロファージのサブタイプは実はとても多様であり,疾患それぞれに深く関連するマクロファージが存在するという考え方.機能も限定的であれば,創薬の標的となりうる.
前述したようにこれまでマクロファージは1種類しかなく,それがM1やM2の状態を行き来すると考えられているが,最近ではマクロファージの亜種(サブタイプ)についても報告がされてきている(概念図).
例えば,CD169+CD11c+マクロファージはリンパ節に常在しており,死んだがん細胞を貪食することにより,がん由来の抗原をCD8T+細胞に直接クロスプレゼンテーションすることによりがんを排除する機能を果たしていることを田中らは報告した14).さらに甲殻類の甲羅の成分でアレルゲンとしても知られるキチンによって活性化されるマクロファージにはJmjd3がその分化に重要であること15),また,マクロファージでありながら,顆粒球のように2核様の特徴的な形態を示すsegregated nucleus containing atypical monocyte(SatM)は,線維化がはじまる際に患部に集積し,線維化の発症に重要であることがわかっている10).その他にも,肥満の過程のなかで死にゆく脂肪細胞から出されたMCP-1によってCCR2+のマクロファージが脂肪周辺をとり囲んで貪食する(crown-like structure)現象も報告されているほか16)17),このマクロファージから分泌される炎症性サイトカインが脂肪分解を促進することで,悪循環を形成することがわかっている18).脳梗塞の際に発生する炎症惹起因子を排除するマクロファージがMafbによって制御されているという報告もある19).
サブタイプとは少し毛色が異なるが,組織のマクロファージの分化や活性化についても研究が進んでおり,それぞれの臓器の環境の影響を受け,それらの機能も多様性に富んでいる.例えば,腹腔内に存在しているマクロファージは,腹膜の一部である体網からのレチノイン酸によって
4本特集の全体像
本特集では,最近のトピックスとして,組織あるいは生命現象や疾患,分子に着目して,マクロファージの最先端をご紹介いただいた.皮膚は外界と触れる器官であり,病原体からの生体防御の観点からだけでなく,アレルギーや感染症,がん等の免疫がかかわる疾患にも重要である.そこで“皮膚とマクロファージ”と題して接触性皮膚炎のマクロファージの役割について,この器官にもともといるマクロファージ,刺激によって誘導されるマクロファージについて解説いただいた(中溝らの稿).病は気からという言葉があるように神経系は免疫系と深く関係しており,興味深い分野である.そのなかでも“痛みとマクロファージ”に焦点をあて,神経系がダメージを受けた後に痛みが発症するという慢性疼痛システムにおいて,神経系にどのように複数のマクロファージが作用するかを寄稿していただいた(津田の稿).また,神経系でも神経変性疾患に着目し“神経変性疾患とマクロファージ”にフォーカスして,普段は中枢神経の保護的作用をもっている神経のマクロファージが,アルツハイマーや筋萎縮性側索硬化症などの神経疾患の発生に伴って異なる活性化型のマクロファージが出現し,それらの割合が病態にかかわることを紹介いただいた(祖父江らの稿).免疫系では腸内環境というテーマは最近の重要なトピックの1つである.“腸とマクロファージ”と題して,その腸での炎症制御とマクロファージの関係性や,実際に腸で起こる疾患として炎症性腸疾患をとり上げた(香山・竹田の稿).前述したとおり,リンパ節内にいるCD169+マクロファージは,放射線照射などによって死んだがん細胞を捕食し,細胞障害性T細胞を活性化することにより抗がん作用を示すことがわかっていた.しかし,最近CD169をマーカーとしてもつ新しい機能について,腸管内に存在しているCD169+マクロファージはケモカインを産生することにより腸内環境の炎症を制御しているという研究が報告された.このようにCD169を切り口としたマクロファージ研究を紹介いただいた(菊池らの稿).線維化は臓器が硬くなり機能しなくなる恐ろしい病気であるが,有効な治療薬はいまだ開発されていない.そこで“線維症とマクロファージ”に焦点をあて,その線維化の発症に重要な疾患特異的マクロファージのサブタイプについてまとめた(佐藤の稿).さらに最近,脂質がマクロファージの活性化と疾患発症に深く関与していることが明らかとなってきている.そこで,“脂質とマクロファージ”に着目して,その脂質代謝と疾患との関係性に関しても記載していただいた(青木・有田の稿).
おわりに
今回は,現時点でのマクロファージについての特集を“疾患”にフォーカスして組んだ.免疫学は日本が世界でも大きな成果を上げている分野の1つであり,日本発のマクロファージ学は世界でトップクラスの素晴らしい研究レベルを保っていると私は考えている.しかし,まだまだ未解明の問題点は山積している.その一つとして,マウスとヒトとの抗原の違いが挙げられる.T細胞やB細胞はマウスとヒトとでは共通のマーカーが存在しているために,マウスでの成果をある程度はヒトに応用することができる.しかし,マクロファージにおいては共通のマーカーはかなり少なく,マーカーがあったとしてもマウスとヒトとではその抗原のもつ生理的意味が違うことがしばしばある.したがって,ヒトにおける創薬への応用を考えるためには,マウスでの解析に加えて,ヒトのマクロファージサブタイプの研究が重要な意味をなす.日本でのマクロファージの基礎研究が応用されて,ヒトにおける研究が進めば,マウスとヒトのマクロファージの種類を俯瞰した地図,いわば日本版マクロファージアトラスが描かれ,その地図をもとにした創薬はこれまでの概念を“破”った新しい作用機序をもった薬の誕生へとつながると期待される.
文献
- 1) Metchnikoff E:Leçons sur la pathologie comparée de l'inflammation. (Lectures on the Comparative Pathology of Inflammation),1892
- 2) van Furth R & Cohn ZA:J Exp Med, 128:415-435, 1968
- 3) van Furth R, et al:Bull World Health Organ, 46:845-852, 1972
- 4) Schulz C, et al:Science, 336:86-90, 2012
- 5) Hoeffel G, et al:J Exp Med, 209:1167-1181, 2012
- 6) Yona S, et al:Immunity, 38:79-91, 2013
- 7) Auffray C, et al:J Exp Med, 206:595-606, 2009
- 8) Hettinger J, et al:Nat Immunol, 14:821-830, 2013
- 9) Kawamura S, et al:Immunity, 46:835-848.e4, 2017
- 10) Satoh T, et al:Nature, 541:96-101, 2017
- 11) Stein M, et al:J Exp Med, 176:287-292, 1992
- 12) Mills CD, et al:J Immunol, 164:6166-6173, 2000
- 13) Gordon S:Nat Rev Immunol, 3:23-35, 2003
- 14) Asano K, et al:Immunity, 34:85-95, 2011
- 15) Satoh T, et al:Nat Immunol, 11:936-944, 2010
- 16) Lumeng CN, et al:J Clin Invest, 117:175-184, 2007
- 17) Cinti S, et al:J Lipid Res, 46:2347-2355, 2005
- 18) Suganami T, et al:Arterioscler Thromb Vasc Biol, 25:2062-2068, 2005
- 19) Shichita T, et al:Nat Med, 23:723-732, 2017
- 20) Okabe Y & Medzhitov R:Cell, 157:832-844, 2014
- 21) Kohyama M, et al:Nature, 457:318-321, 2009
- 22) Satoh T, et al:Nature, 495:524-528, 2013
- 23) Fujiu K, et al:Nat Med, 23:611-622, 2017
著者プロフィール
佐藤 荘:2009年大阪大学医学系研究科博士課程修了(医学/指導教官:審良静男).以降,大阪大学微生物病研究所自然免疫学にてマクロファージの研究を継続しております.今後は得られた基礎研究をもとに“疾患特異的マクロファージ”を標的とした創薬につながるように研究を展開していきたいと思っています.
マクロファージのサブタイプは実はとても多様であり,疾患それぞれに深く関連するマクロファージが存在するという考え方.機能も限定的であれば,創薬の標的となりうる.









