概論
オルガノイド研究を支える発生パターニング
生命進化から俯瞰するパターン形成の原理
Embryonic patterning, a code making organoids research possible
髙里 実
Minoru Takasato:RIKEN Center for Biosystems Dynamics
Research(理化学研究所 生命機能科学研究センター)
オルガノイドは多能性幹細胞や組織幹細胞から形成される臓器様組織であるが,その作製過程は,ヒト個体発生における臓器形成過程,すなわち「パターニング」を順に再現していく作業である.ヒト個体発生には多細胞動物の系統発生の痕跡がパターニングとして残されており,動物がパターニングを獲得してきた原理を考えることで,初期胚から三胚葉への分化,空間的位置情報に基づいた臓器発生,を体系的に理解することができる.本稿では,オルガノイド研究の根底を支える個体発生のパターニングについて,動物の系統発生をなぞって解説する.
はじめに
近代生物学の発展に伴い,モデル動物の解析から膨大な発生生物学的知見が蓄積され,それら知見と近年の幹細胞の培養・操作手法の技術的進歩とが「融合」した結果,「オルガノイド」とよばれる人工三次元器官が生み出された.オルガノイドは,その語源(organ + oid)のごとく,臓器様組織を意味する.これまでに数多くの臓器を対象としたオルガノイドが開発されており,そのなかから創薬や再生医療につながる成果も出つつある.現在,臓器や器官を対象とした新規オルガノイドの開発,オルガノイドの組織構造・機能の質的向上をめざす研究がさかんに行われており,今後も絶え間なく続いていくだろう.オルガノイドは試験管内で多能性幹細胞や組織幹細胞から形成されるが,その過程は,前述の「融合」のくり返しであり,本質的には,個体発生の過程におけるパターニングの原理を見つけ,それをオルガノイド作製に還元していく作業である.発生生物学における「パターニング」とは,胎児内の相対的空間位置に基づいて細胞が異なる種類へと分化する過程,と定義される.多細胞動物が受精卵から個体を発生させる際には,くり返しこのパターニングが行われ,それにより器官や臓器が発生し,さらに体内の正しい位置に正しい向きで配置される.したがって,オルガノイドの開発研究では,個体発生のパターニングを発見し理解する作業からはじめなければならない.
1生物進化に見るパターニングの獲得
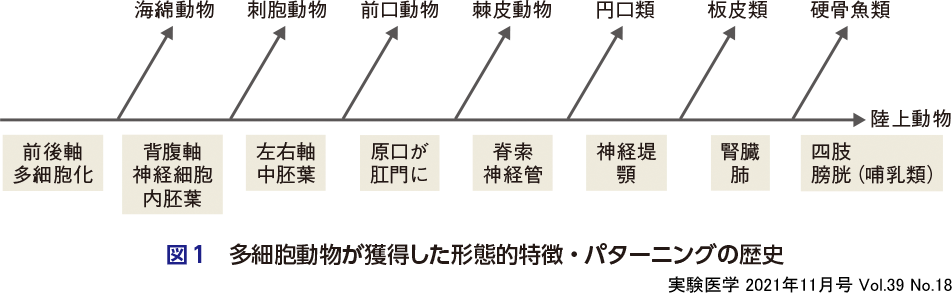
例えば,多能性幹細胞(ES/iPS細胞)から腎臓オルガノイドを作るには,胎児における腎臓の発生過程を試験管内で再現しなければならないが,胎児のなかでは一体どのようにして腎臓は形作られているのだろうか.一般にはあまり知られていないことだが,ヒト胎児の成長過程では,前腎,中腎,後腎という3種類の腎臓が体の前方から後方へ向かって順に発生する.最初に発生する前腎は,中腎の前端に位置する極小の腎臓と考えてよい.次に発生する中腎は,後腎よりも小さい腎臓であり,中腎の一部の管は精巣や卵巣の組織へと変化してゆくものの,最終的には前腎と同様,発生の過程で消失してしまう.そして,最後に最も尾部に発生する後腎こそがわれわれヒトのもつ“腎臓”となる1).なぜこのようなことが起こるかといえば,脊椎動物の進化の過程で前腎,中腎,後腎の順で腎臓が創生されたからである.つまり,ドイツの生物学者ヘッケルが残した「個体発生は系統発生を反復する」という言葉に象徴されるように,ヒト胎児のなかに生命進化の痕跡がパターニングとして残されていることがわかる.多細胞動物の進化過程では,形態的特徴が変化(新しい器官や臓器を獲得)する背後に,新しいパターニング原理の獲得が潜んでいる.そして,その痕跡を辿ると,個体発生におけるパターニングをより体系的に理解することができる(図1).
2初期胚のパターニング
❶ 前後パターニングの獲得
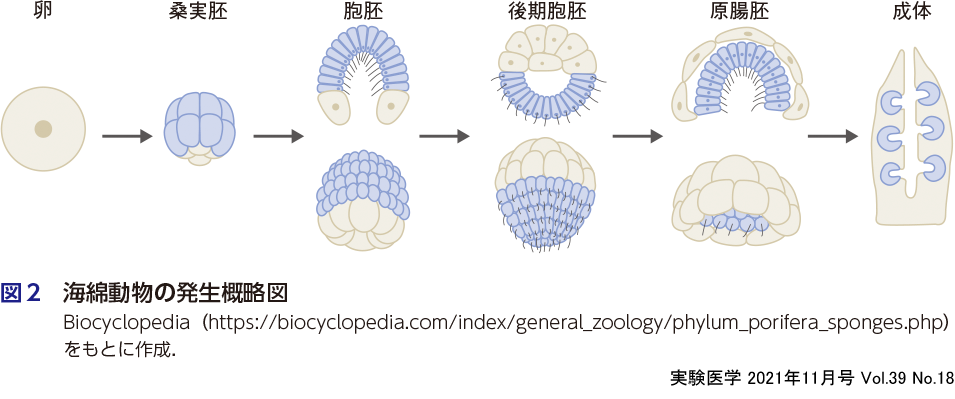
多細胞動物のなかで最も原始的な生物として分類されているのが海綿動物(スポンジ)である.海綿動物は外胚葉しかもたず,外界から体内に水流を取り込み,そこから食物や酸素を得て,老廃物を排出するだけの生物である.その発生過程は非常に単純で,卵細胞から桑実胚(morula),胞胚(blastula),原腸胚(gastrula),幼生(larva)を経由して成長する(図2).多細胞動物が誕生するにあたり,この原腸陥入という発生形態による,食物を効率的に確保するための装置としての「腸」の獲得は必要不可欠な出来事であったに違いない.海綿動物胚発生における最初のパターニングは,原腸胚期に前後軸に沿ってはじまることが知られている.この前後軸の両極に液性因子が(Wntが後極に,TGF-βが前極に)非対称的に発現することで,前後方向にWntとTGF-βの濃度勾配が生まれ,前後のパターニングが生じる2).特に,Wntが発現する側ではβカテニンの安定化による細胞核内移行がみられ(古典的Wntシグナル経路),遺伝子発現が制御されることで,そこが原腸陥入の起点(原口)となる.
❷ 三胚葉の獲得
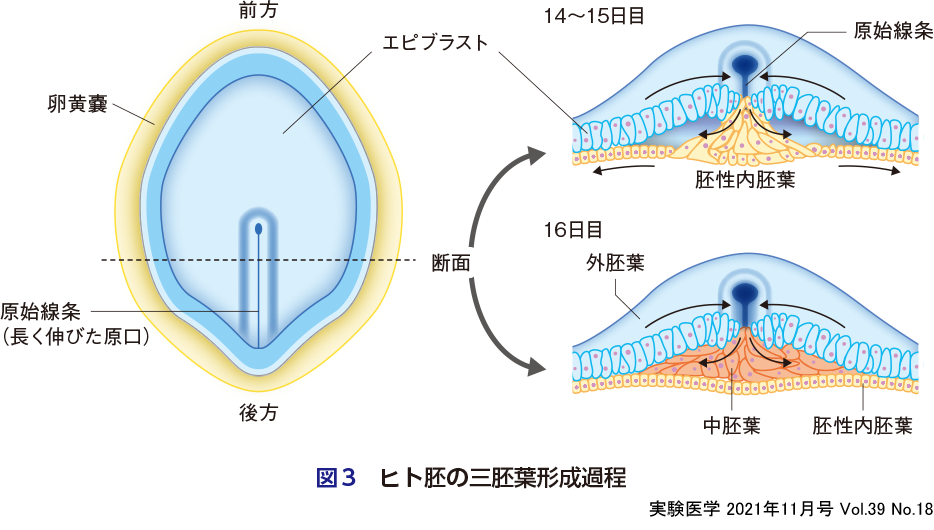
海綿動物からもう1段階複雑化した生物種である刺胞動物(クラゲ・ヒドロ虫など)では,TGF-βの濃度勾配がWntの濃度勾配に対して直角に交差するようになり,前後に加え,背腹方向のパターニングを実現している.その結果,外胚葉と内胚葉(内中胚葉)の二胚葉性が生まれた可能性が考えられている3).多細胞動物が複雑性を獲得するのに,次に重要なステップは中胚葉の獲得である.有櫛動物(クシクラゲ)や,さらに1段階複雑化した左右相称動物になると,前述の内中胚葉が非対称性に分裂し,中胚葉を誕生させる.中胚葉は間葉系細胞であり,上皮細胞である外胚葉と内胚葉の間を充てんする.したがって,中胚葉が発生する際には,原腸胚の上皮系細胞に上皮間葉転換(EMT)が起こるが,これには古典的Wntの下流であるbrachyuryやsnail遺伝子が関与している4).すなわち,原口部分で,予定外胚葉細胞(エピブラスト)から内中胚葉への分化が起こる際に,WntによってEMTが起こった細胞が中胚葉へと分化するというパターニングが生じている.これら原始生物の進化でみられる,三胚葉を生み出すパターニングは,われわれ哺乳類の初期胚においても保存されている(大久保・髙島の稿参照)(図3).例えば,ヒトES/iPS細胞(エピブラストと同質)からの分化誘導系において,Wnt(古典的Wnt)やActivin(TGF-βスーパーファミリー)を添加することで内中胚葉を誘導することができるが,特に古典的Wntを強めることで,βカテニンの核内移行の誘導を経由して,中胚葉を支配的に誘導することができる5).
3三胚葉のパターニング
❶ 外胚葉
外胚葉は根源的には上皮系細胞であり,海綿動物では,外界と胚を隔絶するための表皮としての役割を担っている.その後,刺胞動物や有櫛動物における新たなパターニングの獲得により,その一部が神経細胞(散在神経系)としてはじめて出現した6).外胚葉を表皮たらしめる因子はBmpであり,刺胞動物の原腸胚期では外胚葉で広範囲にBmp2/4が発現している7).一方,原口周辺にはBmpを阻害する因子Chordin,Nogginが発現し,このパターニングにより神経細胞が誕生したと考えられる.実際,ヒトES/iPS細胞から神経組織を誘導する際には,内胚葉と中胚葉への分化を阻害するALK阻害剤(Activin阻害)と,表皮への分化を阻害するTGF-β阻害剤(Bmp阻害)を同時に加える(dual SMAD inhibition法).あるいは,ES/iPS細胞の予定運命が外胚葉であるという性質を利用して,血清抜きの培地で自己組織化的に神経を誘導する方法(SFEBq法:坂口の稿参照)もある.
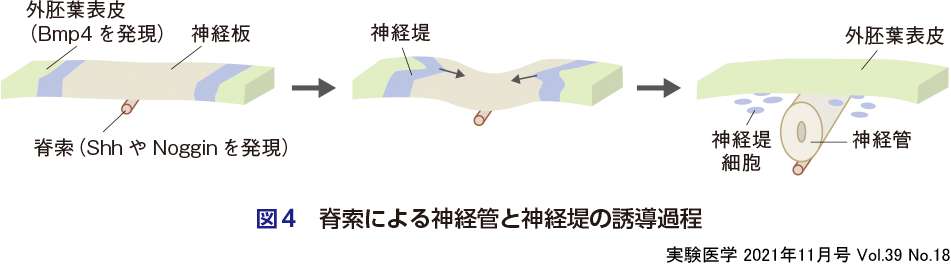
後述する中胚葉由来の「脊索」をもつ脊索動物になると,脊索からのShh(ソニックヘッジホッグ)シグナルにより,外胚葉の一部(神経板)が陥没して,「神経管」とよばれる中枢神経系の源となる神経組織が発生する.このとき,脊索ではBmpを阻害するNogginがShhと同時に発現するが,脊索と反対側の神経板細胞は外胚葉表皮からのBmpの影響を受け,これが神経管の背腹パターニングを決定する(高Bmpが背側,高Shhが腹側の神経管細胞を誘導する)(図4).特に,最も背側の部分(神経管と外胚葉の境界部分)は「神経堤」とよばれる遊走性の細胞となる(図4).神経堤は,第4の胚葉ともよばれるほど特殊な細胞で,全身へ遊走し,末梢神経細胞,メラノサイト,副腎髄質,心臓の平滑筋,顔面表皮,顔面骨格など,さまざまな組織へと分化する.この神経堤の出現が,多細胞動物の形態的複雑化に大きく寄与した.例えば,図1に記載した板皮類以降の生物の「顎」の獲得は,神経堤細胞の出現と同義であり,神経管のパターニングの結果と考えてよいだろう.さらに,神経管の前方部分は脳となり,後方部分は脊髄となる.この中枢神経の前後・背腹軸のパターニングについては,坂口の稿の図2を参照されたい.
❷ 内胚葉
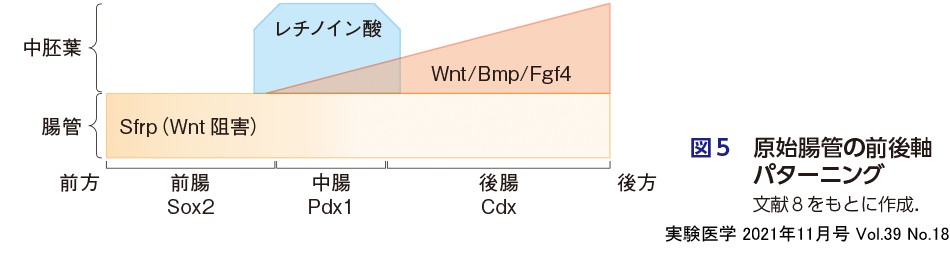
原腸陥入により体の内側を貫通する管状の構造(原始腸管)を形成するのが内胚葉であり,その本質は消化管である.原口が口となるのか,肛門となるのかによって生物種は前口動物(昆虫など)と後口動物(ヒトなど)に分類されるが,いずれにしても前後軸に沿って原始腸管のパターニング(前腸,中腸,後腸)が決定される.この原始腸管の前後のパターニングには,原始腸管とそれをとり巻く中胚葉細胞(臓側中胚葉)との相互作用が重要である.原始腸管上皮はShhを強く発現することで周辺の中胚葉を臓側中胚葉へと分化させるが,逆に臓側中胚葉からのシグナルによって,原始腸管の前後パターンが制御されている.言い換えると,体の前後軸に沿って中胚葉細胞の特徴が変化し,分泌するシグナル因子の種類が変化することが,原始腸管の前後パターニングに影響している.具体的には,後腸は,中胚葉からの高濃度のWnt/Bmp/Fgfモルフォゲンによって誘導され,中腸は中濃度のWnt/Bmp/Fgfモルフォゲンおよびレチノイン酸によって誘導される(船田らの稿参照).一方,前腸では,前腸自身がWntの阻害因子(Sfrp)を分泌することで,Wnt/Bmp/Fgfモルフォゲンをブロックしている(図5)8).
また,原始腸管には,背腹軸のパターニングも存在する.例えば,肺の原芽である気管は,前腸の食道の腹側から発芽する.これは背側の中胚葉がNogginを分泌し,腹側の中胚葉がBmp4とWnt2/2bを分泌することで,気管の発芽に必須なNkx2-1が腹側に発現するというパターニングである(中山・森本の稿参照).また,哺乳類の後腸(総排泄腔)においては,腹側が膨らんで背側から分離されることで膀胱に発達し,背側はそのまま直腸に発達するという背腹軸のパターニングが知られている.
❸ 中胚葉
刺胞動物には中胚葉がないと前述したが,じつは,ヒドロ虫では外胚葉の一部が内胚葉側に膨らみ,Entocodon(いまだ日本語訳はない)とよばれる中胚葉の特徴をもった細胞に分化することが知られている9).Entocodonは,最終的には外胚葉と同様に外界に接する(クラゲの傘の下面を覆う)平滑筋上皮へと分化するが,brachyuryやsnail遺伝子を発現しており,その発生過程は中胚葉の発生過程と似ている.さらに,内胚葉上皮に沿って遊走してきた卵原幹細胞が,この平滑筋上皮細胞層に収まり,そこが生殖腺となる10).すなわち,中胚葉の最も基本的な運命は平滑筋細胞と生殖腺である,と言えよう.ヒト胚においても,始原生殖細胞は原腸に沿って遊走して,中胚葉由来の生殖腺にたどり着く.つまり,この生殖細胞の遊走と生殖腺発生の関係は,多細胞動物の進化のごく初期からヒトに至るまで保存された機構であることがわかる.この機構は中胚葉の背腹パターニングに痕跡として残っており,例えば脊椎動物において,体腔上皮(進化上,最初期の中胚葉性上皮組織で,後述する側板中胚葉由来)は腸管からのShhシグナルにより,腸管に近い側が生殖腺へと分化する11).
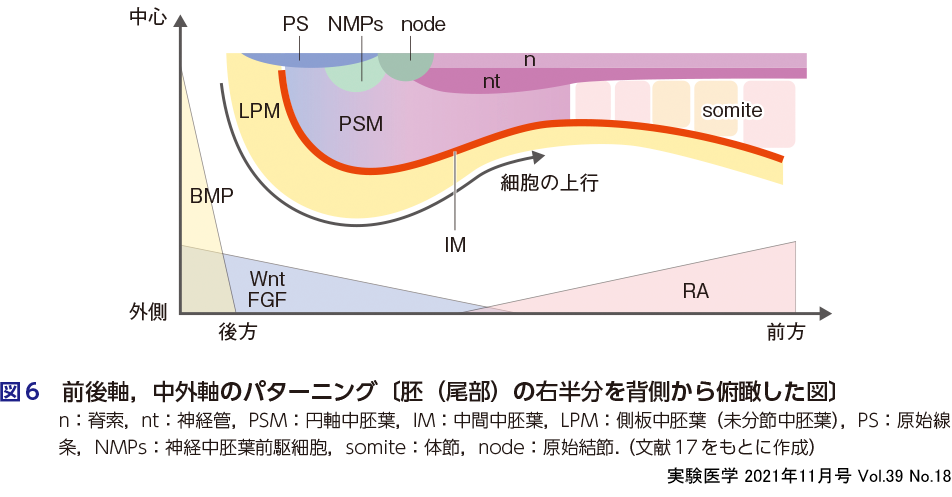
中胚葉が生まれたことで多細胞動物が獲得した重要な要素は脊索の形成である.脊索は脳から尾部まで正中に伸びる棒状の中胚葉組織で,Shhを強く発現し,神経管のパターニングに重要な役割を担う.❶ 外胚葉の項で述べたように,脊索の出現により,中枢神経系の発生,神経堤の発生が導かれた.さらに,脊索を中心として,外側に向かってBmpの濃度勾配が増していき,沿軸中胚葉(骨,骨格筋,血管),中間中胚葉(腎臓),側板中胚葉(平滑筋,生殖腺,間充織,肢芽,心筋)が発生する(図6).
体の前後軸に沿ったパターニングには,ホメオティック遺伝子Hox1〜13が強くかかわっている.Hoxは海綿動物と有櫛動物以外のすべての動物に存在している遺伝子で,胚葉を問わず,頭部から尾部の組織で1から13の順番に発現し,体の前後軸に沿った構造を決定する役割を担っている12)13).例えば,話は遡るが,前腸ではHox1〜4が発現し,後腸ではHox9〜13が発現している8).中間中胚葉由来の腎臓では,腎臓の上皮性前駆細胞であるウォルフ管にはHox7/8が発現しており前方の中間中胚葉から発生するが,間葉系前駆細胞である後腎間葉細胞にはHox10/11が発現しており後方の中間中胚葉から発生することがわかっている(小林・西中村の稿参照)14).また,四肢動物の前肢と後肢もHox遺伝子による制御がされている可能性があり,その結果としてTbx5とTbx4遺伝子が,それぞれ前肢と後肢を規定している.この中胚葉の前後軸パターニングには,初期胚のある一時に体幹部分に特異的に発現するレチノイン酸がかかわっている(森・永樂の稿参照)12).なお中胚葉の発生では,原口(原始線条)でエピブラストから分化した中胚葉が,尾部から体幹部へと上行していく.したがって,中胚葉の前後軸パターニングは,レチノイン酸などのモルフォゲンの濃度勾配と,細胞が上行するタイミングとの組合わせにより,時空間的に制御される(図6)1)15).
おわりに―オルガノイド技術への応用
パターニングを司る因子や環境を理解し操ることで,ヒト多能性幹細胞の分化誘導を個体発生の枠にはめ,よりヒト生体の臓器に近いオルガノイドの作製が実現できる.そこで,本稿では個体発生のパターニングについて,動物の系統発生をなぞって解説し,その概要の体系的な理解を試みた.そして各論では,各器官について,発生パターニングの解説とオルガノイド作製への応用例を紹介いただく.形態的特徴に基づく伝統的な進化学は,遺伝的分子的エビデンスに基づく進化学へと発展してきたが,今後は1細胞オミクス技術をとり入れた進化学が期待され16),パターニング原理のより精緻な理解が進むだろう.実際,この10年ほどのオルガノイド技術の進歩は目覚ましく,ありとあらゆる臓器のオルガノイドが多能性幹細胞から作製され,その複雑性や再現性も年々高度に進化してきた.一方,三次元的に進行する個体発生のパターニングが理解できたとして,その原理を,そのまま実験室の細胞培養系に当てはめるには,技術的な課題が立ちはだかっている.例えば,モルフォゲンの三次元的な濃度勾配をどう実現するか,成長するオルガノイドへの酸素・栄養供給,つまり血管化と血液供給をどのように行うか,などは特に重要な問題である.本特集の各論でも述べられているように,多くのオルガノイド研究者は医工学的なアプローチの重要性を認識している.次の10年は,その課題解決の進展に期待したい.
文献
- Takasato M & Little MH:Development, 142:1937-1947, doi:10.1242/dev.104802(2015)
- Adamska M, et al:PLoS One, 2:e1031, doi:10.1371/journal.pone.0001031(2007)
- Martindale MQ:Nat Rev Genet, 6:917-927, doi:10.1038/nrg1725(2005)
- Salinas-Saavedra M, et al:Elife, 7:doi:10.7554/eLife.36740(2018)
- Gadue P, et al:Proc Natl Acad Sci U S A, 103:16806-16811, doi:10.1073/pnas.0603916103(2006)
- 小泉 修:比較生理生化学,33:116-125,doi: 10.3330/hikakuseiriseika.33.116(2016)
- Hayward DC, et al:Proc Natl Acad Sci U S A, 99:8106-8111, doi:10.1073/pnas.112021499(2002)
- Zorn AM & Wells JM:Annu Rev Cell Dev Biol, 25:221-251, doi:10.1146/annurev.cellbio.042308.113344(2009)
- Spring J, et al:Dev Biol, 244:372-384, doi:10.1006/dbio.2002.0616(2002)
- Seipel K, et al:Dev Dyn, 230:392-402, doi:10.1002/dvdy.20061(2004)
- Yoshino T, et al:Nat Commun, 7:12561, doi:10.1038/ncomms12561(2016)
- Deschamps J & van Nes J:Development, 132:2931-2942, doi:10.1242/dev.01897(2005)
- DuBuc TQ, et al:Nat Commun, 9:2007, doi:10.1038/s41467-018-04184-x(2018)
- Brunskill EW, et al:Dev Cell, 15:781-791, doi:10.1016/j.devcel.2008.09.007(2008)
- Takasato M, et al:Nature, 526:564-568, doi:10.1038/nature15695(2015)
- Siebert S, et al:Science, 365:doi:10.1126/science.aav9314(2019)
- Tani S, et al:Exp Mol Med, 52:1166-1177, doi:10.1038/s12276-020-0482-1(2020)
著者プロフィール
髙里 実:東京大学大学院理学系研究科生物科学専攻(浅島誠先生)および東京大学医科学研究所(西中村隆一先生)に師事し,マウス腎臓発生の研究を行い2008年に博士号(理学)取得.その後,オーストラリア・クイーンズランド大学(Melissa Little教授)へ留学し,ヒトES細胞から腎臓オルガノイドを作製した.現在は,腎臓を中心とした泌尿器系臓器オルガノイドをヒト多能性幹細胞から作製する研究,および発生学研究を推進.
 腎臓から眺める生命進化
腎臓から眺める生命進化










