
概論
TRPチャネル研究の現在と未来
TRP channel research: from the resent to the future
富永真琴
Makoto Tominaga:Division of Cell Signaling, Okazaki Institute for
Integrative Biosciences (National Institute for Physiological
Sciences)〔岡崎統合バイオサイエンスセンター(生理学研究所)
細胞生理研究部門〕
TRPチャネルは,1989年にショウジョウバエのtrp遺伝子が同定されて以来,世界で精力的に研究されてきた.Ca2+透過性の高い非選択性陽イオンチャネルとして多彩な細胞機能にかかわることが明らかとなり,チャネル病も数多く,さまざまな後天的疾患やがんの発生にかかわることもわかっている.今後,TRPチャネルの構造や機能の解析が進むことによって,Ca2+やNa+流入にかかわる細胞機能が明らかになり,TRPチャネル機能や発現を制御することで疾患を治療できるようになることを期待したい.TRPチャネルは今もっとも注目を集める分子群の1つと言えよう.
はじめに
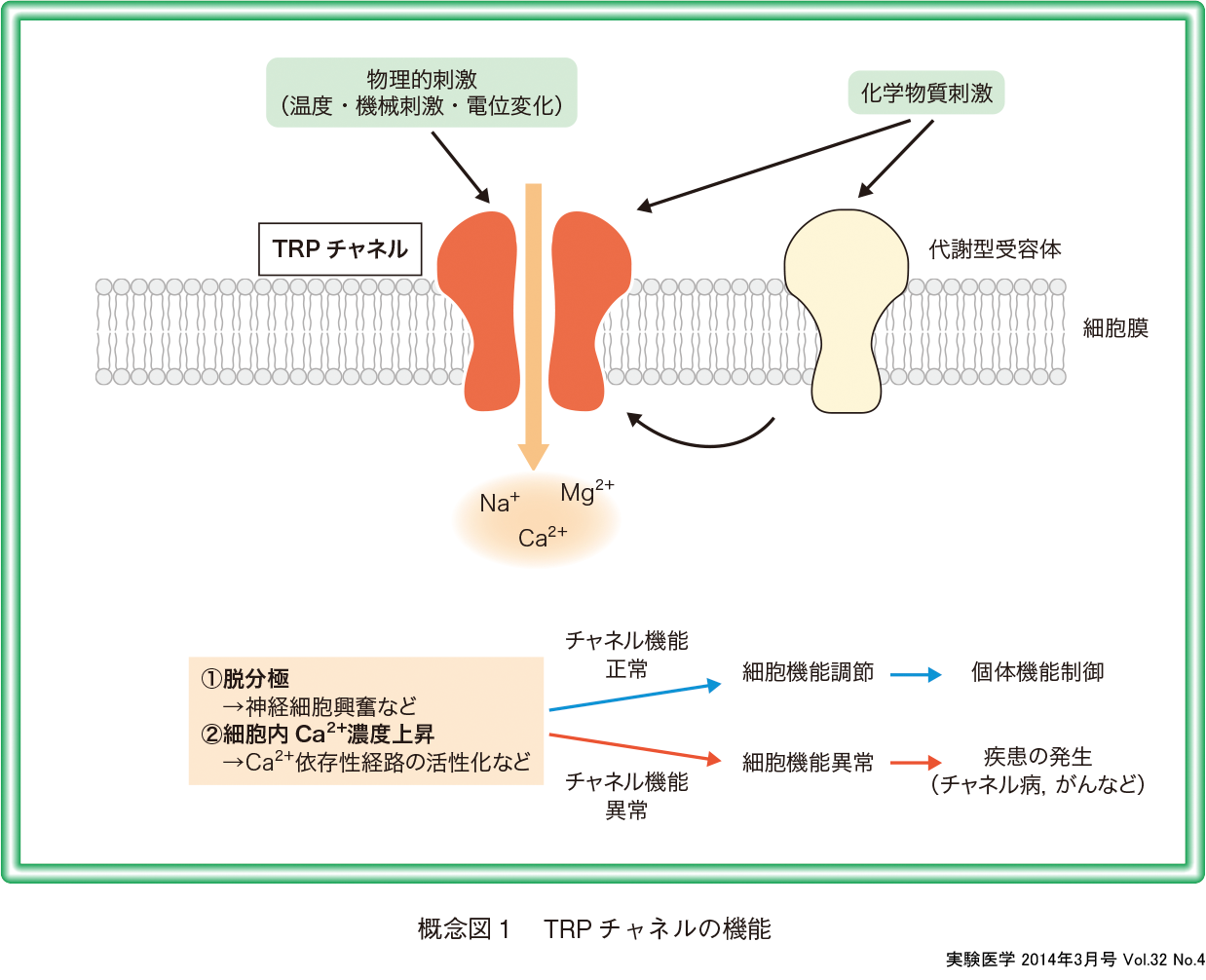
TRP(transient receptor potential; “トリップ” とよばれる)チャネルは,1989年にショウジョウバエのtrp遺伝子が同定されて以来,世界で精力的に研究され,大きな機能的多様性を有するイオンチャネルファミリーを形成することが示されてきた.それまで「非選択性陽イオンチャネル」(non-selective cation channels)と電気生理学的に括られていた一群のチャネルの分子実体が明らかになったことで,その生理的意義が細胞,種を越えて議論できるようになったのである.Ca2+透過性が高いためにチャネル開口によるCa2+流入が細胞内のさまざまなCa2+依存性経路を活性化し,神経細胞においては脱分極から細胞興奮をもたらす (概念図1).このようなTRPチャネルの発見の歴史とこれからの研究の方向を議論したい.
1TRPチャネルの分類と歴史―発見から25年
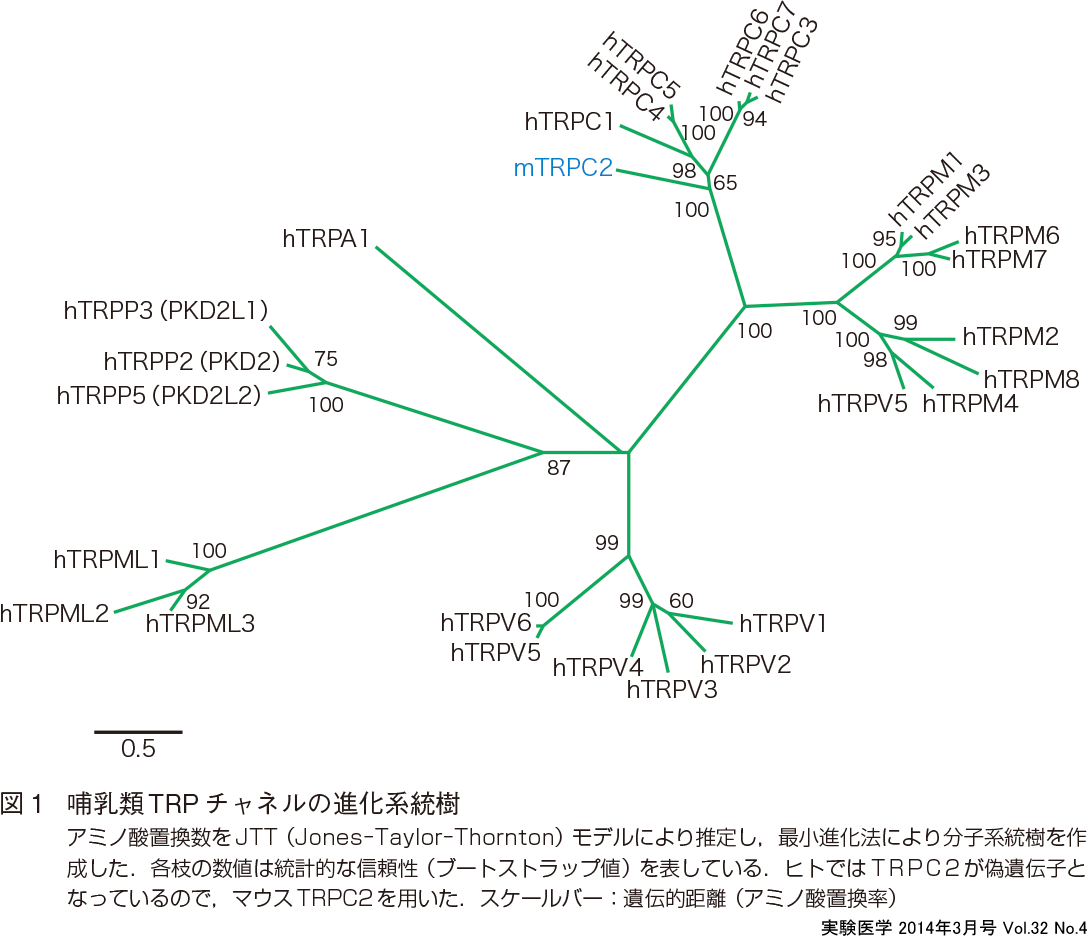
trp遺伝子は1989年にショウジョウバエの光受容応答変異株の原因遺伝子として発見され た1).trp変異株において光刺激に対する受容器電位 (receptor potential) 変化が一過性 (transient) であることから命名された.trpがコードするタンパク質 (TRP)の多くは非選択性陽イオンチャネルを形成している.2002年にTRPC,TRPV,TRPMサブファミリーについて分類と名前の統一が行われた2).遺伝子解析の結果,図1に示すように多くのTRPホモログが同定され,TRPイオンチャネルスーパーファミリーは7つのサブファミリー:TRPC(canonical),TRPM (melastatin),TRPV (vanilloid),TRPML (mucolipin),TRPP (polycystin),TRPA (ankyrin),TRPN (nompC:no mechanoreceptor potential C)に分けられるが,哺乳類にはTRPNサブファミリーはなく28のチャネルが,そしてヒトではTRPC2が偽遺伝子となっており27のチャネルが6つのサブファミリーを構成している3)4).
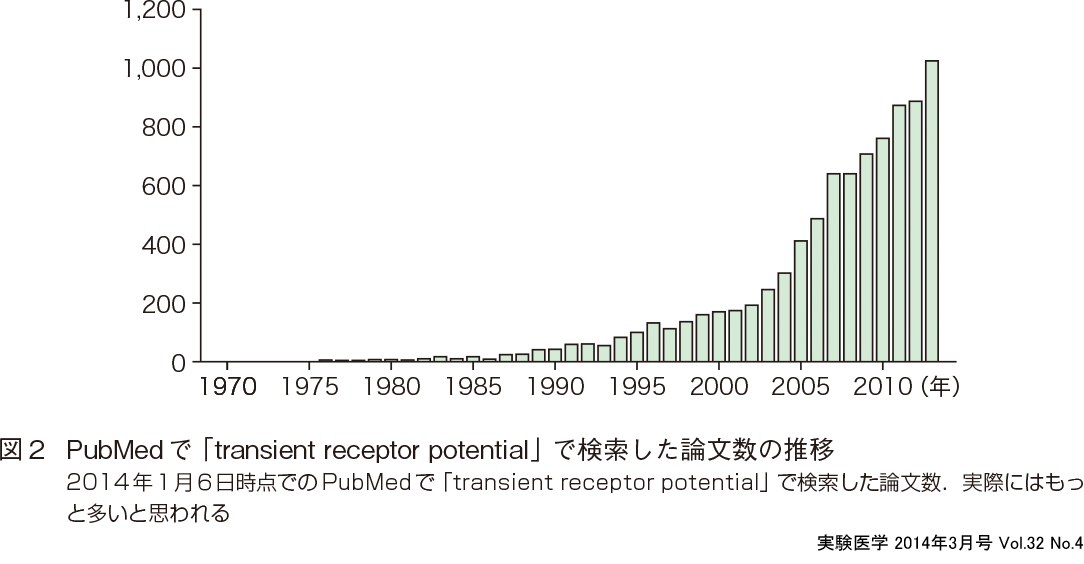
TRPチャネル研究のブレイクスルーは,そのいくつかがセンサーとしてさまざまな細胞外刺激の感知を行うことが明らかになったことである(加塩・富永の稿).その最初の分子がカプサイシン受容体として知られるTRPV1(1997年に遺伝子クローニング)である5).感覚に関与する受容体として研究がもっとも遅れていたのが物理刺激センサーであり,TRPV1は直接熱を感知して活性化することから大きな注目を浴びた.その論文は2014年1月6日の時点で被引用数が3,808回を数える.PubMedで「transient receptor potential」と入力すると約9,000の論文が該当する(図2).TRPV1の遺伝子クローニング論文が3,808回引用されていることを考えると,この数字は実際の論文数よりかなり少ないと思われるが,それでも,ここ数年での指数関数的な論文数の増加が見て取れる.また,TRPチャネルが直接センサーとして機能することに加え,主にTRPCチャネルが受容体作動性チャネルとしてさまざまな代謝型受容体の下流で活性化することも研究の進展に寄与した.電位作動性Ca2+チャネル研究者の多くがCa2+透過性の高いTRPチャネル研究をはじめたことも関係しているかもしれない.
2TRPチャネルの構造―ついに見えた巨大膜タンパク質の姿
TRPチャネルの一般的な構造としては,1つのサブユニットが6回の膜貫通ドメイン(セグメント)を有し,第5,第6膜貫通ドメインの間にイオンを通すと考えられる短い疎水性領域が存在し,N末端,C末端とも細胞内にあると推定されている.四量体で機能的なチャネルを形成すると予想されている.TRPC,TRPV,TRPAサブファミリーではN末端にアンキリンリピートドメインがあり,細胞質内分子との結合を含むさまざまなチャネル機能制御機構にかかわっていると考えられている.第6膜貫通ドメインのすぐ後のC末端細胞質内領域には “EWKFAR”というTRPボックスを含む25アミノ酸からなるTRPドメインが存在する.このTRPドメインやTRPボックスに似た配列は,TRPC,TRPM,TRPVサブファミリーにだけみられる.TRPチャネルのいくつか(TRPV1,TRPM8,TRPM4,TRPM5,TRPA1)には電位依存性があることが知られているが,TRPM8を除いて第4膜貫通ドメインに電位作動性チャネルに特徴的な正電荷をもったアミノ酸はなく,電位依存性の構造基盤は不明である.
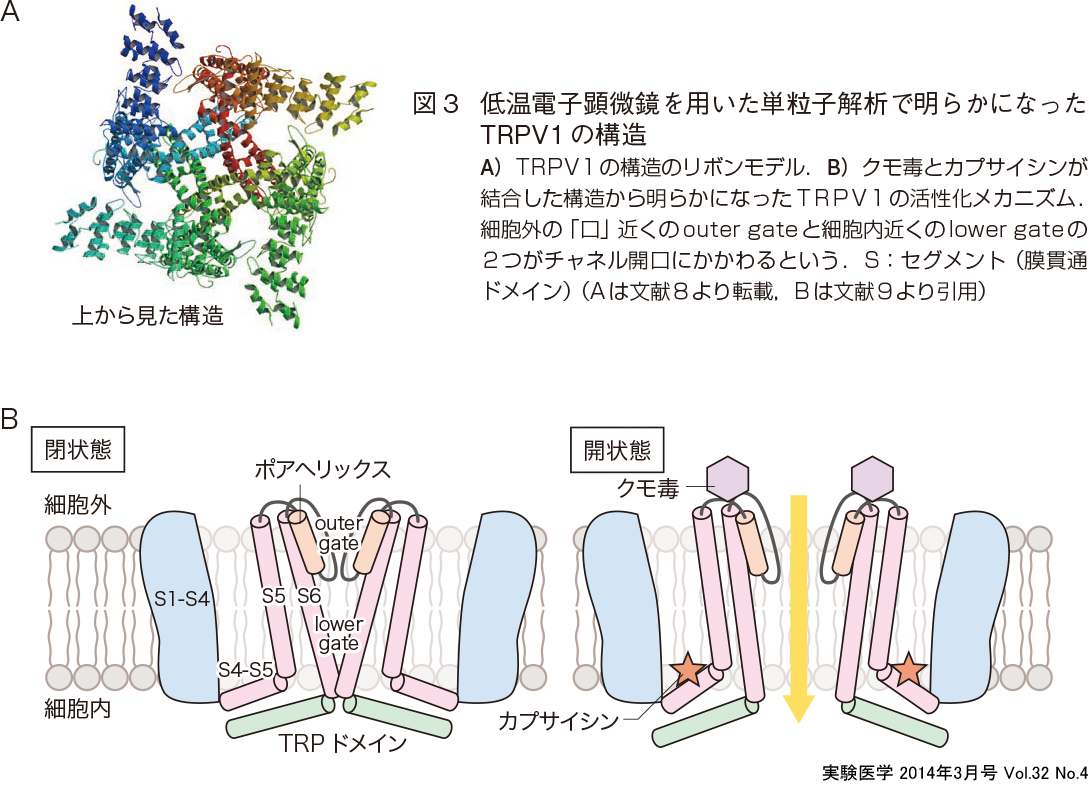
大きな膜タンパク質であるTRPチャネルタンパク質の結晶化は難しく,部分的な結晶構造しか明らかにされていなかった.そのようななか,TRPV1,TRPV4,TRPC3,TRPM2,TRPA1などにおいて,単粒子解析から10〜15Åの解像度でドーム状やバスケット状の構造をもっており,4回対称の四量体であることが明らかになっている(三尾らの稿).加えて,2013年の12月に低温電子顕微鏡を用いた単粒子解析からTRPV1のほぼ全体の構造が3.4Åという高い解像度で明らかにされて大きな注目を浴びている(図3A)6)7).TRPV1は短い「選択性フィルター」(selective filter)と大きな細胞外の「口」(mouth)構造をもち,TRPドメインは第4,第5膜貫通ドメインリンカーと相互作用し,サブユニット構成にN末端のアンキリンリピートドメインが重要な役割を果たすという.明らかにされた原子レベルでの構造をもとに,TRPチャネルの構造と機能の理解がさらに進むものと期待されるし,ほかのTRPチャネルの高解像度の構造解析も同じ手法で行われるようになるであろう.TRPV1のカプサイシンとクモ毒vanillotoxinが結合した状態での構造解析から,outer gate(選択性フィルターを内包する)とlower gateの2つのゲーティング機構をもつことが明らかとなった(図3B)が,未だどのようにして温度刺激がチャネル開口をもたらすかは不明である.
3TRPチャネルを標的とした創薬―構造をもとに高まる期待
TRPチャネルの多くがセンサー機能をもつことから(中川の稿),感覚制御の側面からTRPチャネルは格好の創薬標的になると考えられた.また,Niliusの稿で述べられるように,TRPチャネル機能異常は多くのチャネル病をひき起こし,多くの後天的疾患やがんの発生においてTRPチャネルが重要な役割を果たしていることが明らかにされ(北島・西田の稿,井上・胡の稿)阻害薬あるいは刺激薬の有用性が大いに期待されている.特に,感覚神経細胞のように限られた細胞での限局的な発現が明らかになっているTRPチャネルに対して世界中で作用薬開発が進んだ.カプサイシン受容体TRPV1は複数の侵害刺激受容(痛み感覚) にかかわることから,その阻害薬はよい鎮痛薬になると期待されたが,遺伝子クローニングから17年,未だ市場に阻害薬は現れていない10).いくつかの薬剤が第二相臨床試験に進んでいるとされるが,論文としてヒトでの痛みに関する効果(熱性痛みの閾値の上昇)が報告されているのは少なく,ドロップアウトしたとされる化合物でも鎮痛効果に関してはほとんど言及されていない.大きな副作用として体温上昇や温度感覚異常が知られている.明らかになった原子レベルでの構造をもとにTRPV1作用薬開発が今後,急速に進展するものと期待されるが,ほかのTRPチャネルも構造をもとに創薬が進むことが予想される.
4TRPチャネルの進化―多様な環境に適応するために
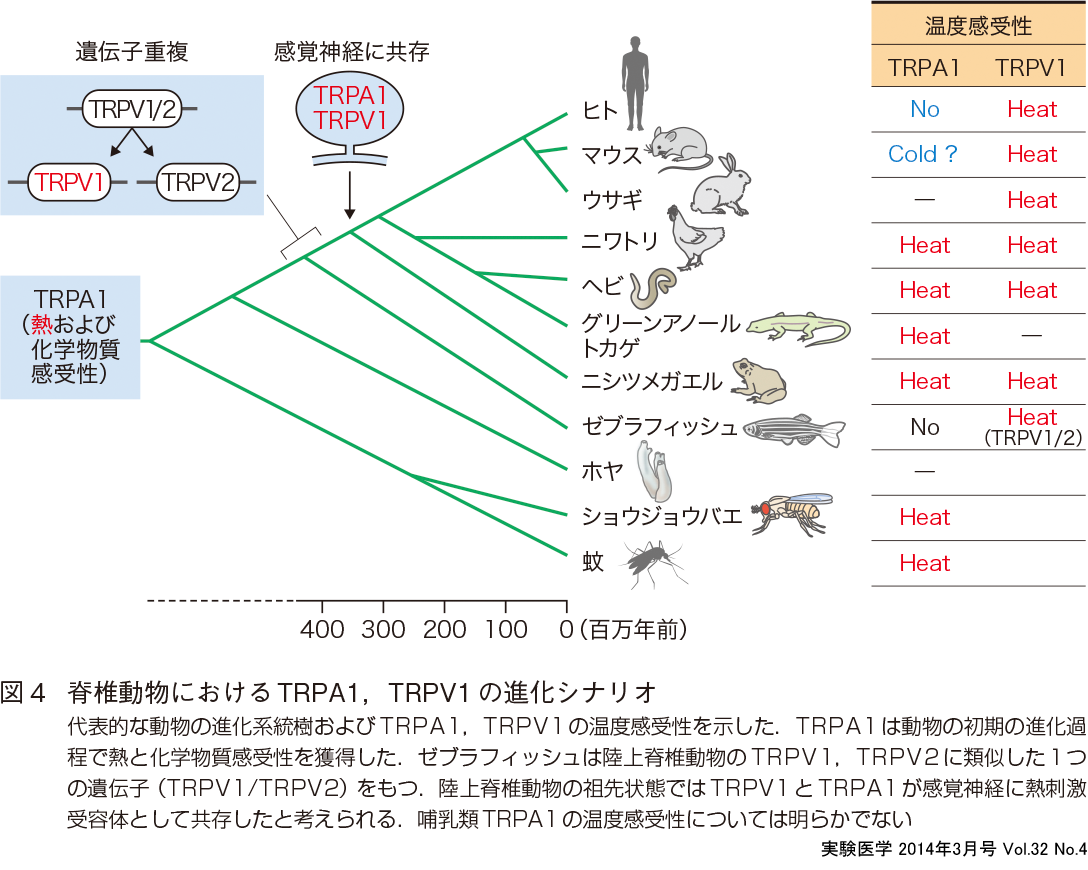
最後に,TRPチャネルファミリーの進化についてふれる.TRPチャネルがセンサー機能をもっていることは,生物が進化の過程でセンサー機能を変化させて環境変化に適応してきたことを予想させる.また,線虫や昆虫もTRPチャネルを感覚に利用しており,このことは,TRPチャネルが祖先種から存在する重要なタンパク質であることを意味する.しかし,ショウジョウバエは哺乳類とは異なる種類のTRPチャネルを温度受容体として用いていることが分子系統解析により示されており11),温度受容体は進化過程で種類や機能を柔軟に変化させてきたことがわかる.温度感受性TRPチャネルに焦点をあてて分子系統解析を行うと,ヒトや齧歯類の温度感受性TRPチャネルホモログのほとんどは硬骨魚類と陸上脊椎動物の祖先種にはすでに備わっていたことがわかった.その後に,遺伝子重複や遺伝子欠失がそれぞれの進化系統で独自に生じた結果,遺伝子のレパートリーが脊椎動物種間で多様化してきたことが明らかとなっている.侵害刺激受容体として機能するTRPV1,TRPA1に絞ると,昆虫は複数種のTRPAチャネルをもっており,そのいくつかが熱刺激で活性化する.一方,TRPV1は魚類で現れることから,TRPA1の方が進化的に古いと言える.TRPA1は齧歯類では冷刺激感受性であり,ヒトでは温度感受性がないと理解されており,進化の過程で温度感受性が変化したことになる.脊椎動物で進化系統樹に沿ってTRPA1の温度感受性を解析すると,両生類,爬虫類ではTRPA1は熱刺激センサーとして機能しTRPV1と近い活性化温度閾値を有する12).驚くべきことに,ニワトリTRPA1も熱感受性であることがわかり,恒温動物と変温動物の境界はTRPA1の熱感受性の境界とは異なっていた(図4)13).おそらく,哺乳類への進化の過程でTRPA1の熱感受性が失われたのであろう.ヘビでは,ピット器官とよばれる赤外線感知器管をもつ毒蛇ほど三叉神経に発現するTRPA1の活性化温度閾値が低いという.獲物である哺乳類を感知しやすいようにTRPA1の温度感受性を変化させたものと理解される.このように,生物の進化と関連させたTRPチャネルの解析は,TRPチャネルの生理機能を理解するうえで必要不可欠である.
おわりに
1989年にtrp遺伝子が報告されてから25年,世界中の研究者がこの「非選択性陽イオンチャネル」の解析を進め,多くのことが明らかになってきた.しかし,Ca2+透過性の高い「非選択性陽イオンチャネル」は非常に多くの細胞機能にかかわると想像され,まだ,ほんの少ししか明らかにされていない.1日も早い全容解明が望まれる.そして,TRPチャネルを制御することが,細胞機能,組織機能を,ひいては個体機能を制御することにつながることが理解され,原子レベルでの構造解明の上にTRPチャネルを標的とした薬剤が開発されることが期待される(概念図2).TRPチャネルと疾患との関連ももっと研究されていくであろう.シミュレーション解析も進むものと期待される(井上・胡の稿).しかし,TRPV1の構造が解かれた今でも,どのようにして温度刺激がチャネル開口をもたらすかは明らかでなく,温度などの物理刺激によってTRPチャネルが開口するダイナミックな構造変化を解析可能な技術の開発が大いに望まれる.
本特集を通じてTRPチャネル研究の「今」とおもしろさをお伝えすることができれば,これに勝る喜びはない.
謝辞
分子系統樹作成に関して,岡崎統合バイオサイエンスセンター(生理学研究所)細胞生理研究部門 齋藤 茂 博士に深謝したい.
文献
- Montell, C. & Rubin, G. M.:Neuron, 2:1313-1323, 1989
- Montell, C. et al.:Mol. Cell, 9:229-231, 2002
- 沼田朋大 ほか:生化学,81:962-983, 2009
- Gees, M. et al.:Compr. Physiol., 2:563-608, 2012
- Caterina, M. J. et al.:Nature, 389:816-824, 1997
- Liao, M. et al.:Nature, 504:107-112, 2013
- Cao, E. et al.:Nature, 504:113-118, 2013
- Liao, M. et al.:RCSB Protein Data Bank, PDB ID:3J5P
- Henderson, R.:Nature, 504:93-94, 2013
- Moran, M. M. et al.:Nat. Rev. Drug Discov., 10:601-620, 2011
- 齋藤 茂:比較生理生化学,28:259-266, 2011
- Saito, S. et al.:J. Biol. Chem., 287:30743-30754, 2012
- Saito, S. et al.:Mol. Biol. Evol., in press(2014)
参考図書
- 「TRPチャネルの構造と多様な機能」(沼田朋大 ほか),生化学,81:962-983, 2009
- 「温度感受性TRPチャネルと疾患」(富永真琴),医学のあゆみ,245:831-837, 2013
著者プロフィール
富永真琴:1984年,愛媛大学医学部医学科卒業,臨床研修の後,’92年に京都大学大学院医学研究科博士課程を修了し,生理学研究所助手となる.カリフォルニア大学サンフランシスコ校でTRPV1の解析にかかわってから温度感受性TRPチャネルを研究している.2000年,三重大学医学部教授,’04年から現職 (岡崎統合バイオサイエンスセンター教授).温度がかかわる生命現象をTRPチャネルに焦点をあてて解明したいと思っている.
 温度センサーTRPV1発見秘話
温度センサーTRPV1発見秘話









