
概論
古くて新しい,小さくて大きい細胞の「毛」
A hair of cells that is old but new, and small but big
井上尊生
Takanari Inoue:Department of Cell Biology, Johns Hopkins University(ジョンズホプキンス大学細胞生物学分野)
一次繊毛は細胞に生えた「毛」のように見える構造で,1世紀以上前の文献にすでに記述がある.しかし長い間科学者達は,一次繊毛が細菌の鞭毛などの痕跡器官ではないかと考えていた.一次繊毛は直径が約250 nmと光の回折限界に近いこと,1つの細胞に1つしかないこと,また他の細胞内小器官と異なり脂質膜で区切られていないために通常の生化学的・細胞生物学的実験の適用が困難なことも研究の障害となってきた.近年になって,一次繊毛の構造と機能がさまざまな遺伝疾患と強い結びつきがあることがわかり,研究が爆発的に進んできておりその結果,一次繊毛がわれわれの体のなかで非常に重要な役割を担っていることもわかってきた.最近では,さまざまな科学分野の技術や知識が一次繊毛の研究に集約され,この小さな「毛」のなかで一体何がおきているのかを可視化し,さらに操作することで,その機能を詳細に解明することが試みられている.これらの研究成果の一環として,比較的静的だと思われてきた一次繊毛が,時には動的なふるまいをすることもわかってきた.しかしながら,一次繊毛の構造や機能に関する分子レベルでの理解は端緒についたばかりである.本特集では,この古くて新しい一次繊毛の現時点での構造と機能,そして病態に関する知見をまとめ,さらに小さいがゆえの技術課題に特に焦点を当て,概観する.現在までの知識を俯瞰することで,直近および次世代の研究課題をあぶりだすことも期待している.さらにこの特集を通して,領域横断的に知識や技術を結集することの大切さも読者と共有したい.執筆に関しては,この研究歴史の長いオルガネラに挑戦する新しい精鋭達を応援する意味を込めて,比較的若い方を選定させていただいた.彼ら・彼女らのフレッシュで熱いメッセージや筆の勢いを,美しい一次繊毛の写真とともに楽しんでいただけたら幸いに思う.
はじめに
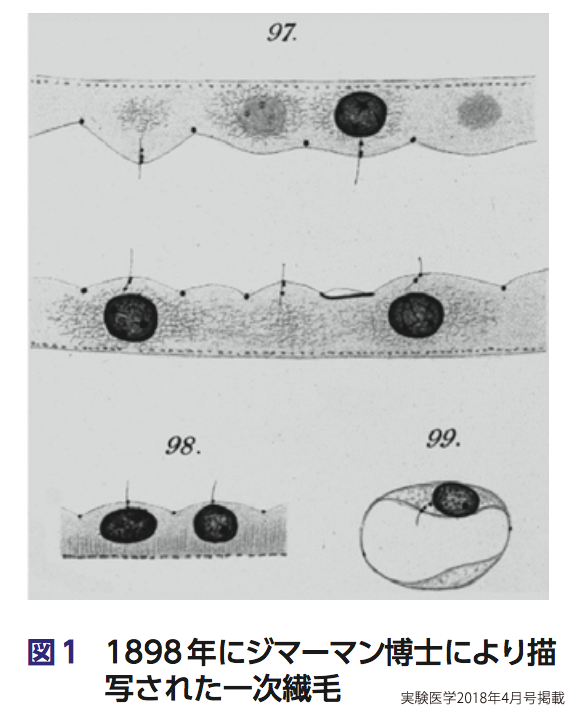
19世紀末,明治維新に端を発するさまざまな変化が日本で起きるなか,ドイツの顕微鏡解剖雑誌に一つの論文が発表された.「上皮と腺に関する知識への貢献」と題されたその論文はスイスの解剖学者カール・ジマーマンによるもので,科学史のなかではじめてヒトを含む哺乳動物細胞の表面に生える「毛」,一次繊毛を描写した(図1)1) 2).そしてこの構造体が,特に腎臓において細胞の知覚にかかわっているのではないかと予想した.しかし多くの研究者は,こうした毛は痕跡器官だと考えた3).単細胞生物の運動を司る繊毛や鞭毛が,多細胞生物となって進化していく過程で当初の役割を失い,構造物としての痕跡だけを残したという考えだ.そうした理由もあって,一次繊毛の機能についてはその後あまり研究が進まなかった.時は過ぎて1968年,ハーバード大学の解剖学者セルゲイ・ソロキンは,肺の上皮細胞の電子顕微鏡像を異なる発生段階で撮像し,細胞に生える毛の順序や種類,形成機構を推測した4).そして一番はじめにはえた毛を一次繊毛とよぼうと提唱した.「一次」は「最初に」の意味だ.つまりソロキンの観察による,肺の上皮細胞は発生段階においてまず一次繊毛が形成され,次の発生段階では1細胞につき複数の運動性を有する繊毛(動繊毛)が形成されることに由来する.さらに時は過ぎ,20世紀終わりから21世紀にかけて,一次繊毛がさまざまな遺伝疾患と深く関連することが遺伝学的研究から明らかとなり,一次繊毛の機能と構造に関する研究がさかんになった.
1構造と盛衰
一次繊毛は,われわれの体を構成する大部分の細胞種で観察されている.一次繊毛が確認されていない細胞種として血球細胞があげられるが,特殊な実験条件下では培養した白血球細胞も一次繊毛を形成することから,形成する能力はあるようだ5).一次繊毛の基本構造を成すのは,中心体を構成する母中心小体に由来する基底小体とそれを足場に伸長した微小管である.動繊毛とは大きく異なり,中心の微小管 (中心対微小管)とそれに付随する構造体やモータータンパク質は一次繊毛には存在しない.一次繊毛の長さは組織によって異なるが,おおむね1〜5 μmである.一方でこれらの法則に従わない特殊な例も存在する.例えば,胚の一部であるノードの,ピット細胞とよばれる細胞は中心対微小管がないにもかかわらず運動性を保持し,また嗅球細胞は運動性がないにもかかわらず中心対微小管を保持し,また一つの細胞につき複数の一次繊毛が生え,さらに長さは20 μmにもなる.こうした亜種は存在するものの,一次繊毛の一般的な定義は,「運動性のない,細胞に一つだけ存在する,基底小体および微小管からなる,細胞表面の突出した構造体」となる.参考までに(図2、本誌参照)に走査型電子顕微鏡で観察した血管内皮細胞の一次繊毛を示す.
一次繊毛の形成と分解は,細胞周期を軸として高度に制御されている(小林の稿,二本垣の稿).また小胞体やミトコンドリアといった他の細胞内小器官と比べてユニークな点は,脂質膜で完全に仕切られていないことがあげられる.一次繊毛の内腔は細胞質と直接つながり,一次繊毛の膜は細胞形質膜と直接つながる.一方で一次繊毛と細胞質の境界面には,移行帯とよばれる特殊なバリア構造が形成される(高尾の稿).このバリアがタンパク質の往来を選択的に制御する.そして一次繊毛内外の能動的な物質輸送は,IFT(intraflagellar transport)とよばれる分子複合体が担う.移行帯とIFTの共同作業により一次繊毛内に特殊な生化学的環境が達成され,それが機能発現の基盤となっている.
2機能と組織と病気
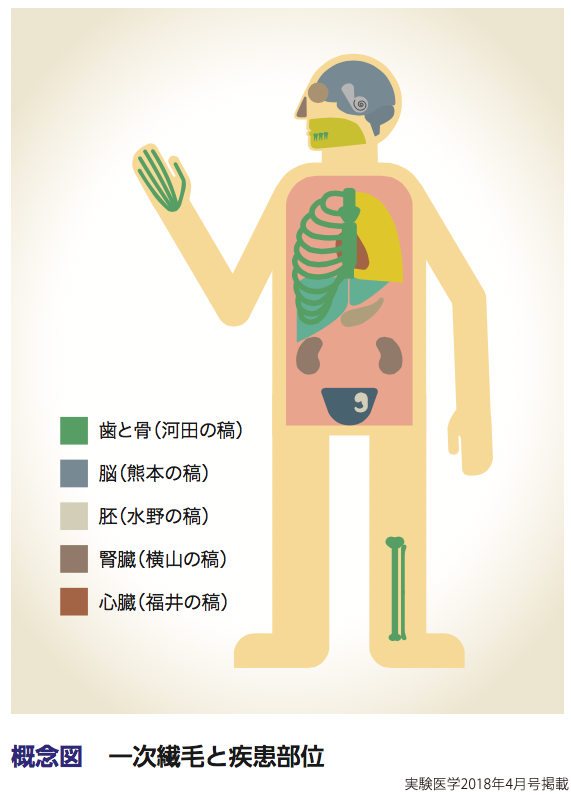
一次繊毛は細胞のアンテナとして機能する(概念図).検出の対象は組織によって異なるが,成長因子,ホルモン,神経ペプチドなどの生化学的な要素から,光,温度,重力,ずり応力などの物理的な要素までと多岐にわたる6).例えば,脳においてはHh(ヘッジホッグ)リガンドやニューロペプチドを検知することで,神経幹細胞や神経細胞の分化,増殖,移動などを制御する(熊本の稿).発生期の心臓では,心拍由来の液体の流れやHhリガンドや増殖因子などを検知することで,心臓の形態形成にかかわっている(福井の稿).ノードとよばれる胚の一部では,周囲の動繊毛が引き起こす液体の流れを感知することで,体の左右軸の決定に重要な役割を担う(水野の稿).また腎臓の管腔においては,原尿が流れることによるずり応力を検知し,細胞分裂軸の決定をする(横山の稿).網膜の桿体細胞の外節は一次繊毛が極度に分化したものだが,ロドプシンを介して光子を知覚する.また歯や骨といった硬組織においても一次繊毛は重要な機能を有する.骨や歯では,Hhリガンドや増殖因子に加えて機械的な刺激を検知するメカノセンサーとしてはたらくことで,軟骨組織や歯牙組織を形つくる細胞の配置を制御している(河田の稿).
一次繊毛がこうしたアンテナとしての機能を発現できる原動力は,一体なんだろうか?その答えに触れる非常に重要な論文が2012年に発表される6).一細胞における一次繊毛の数を実験的に増やすと ,同程度の増殖因子刺激に対する応答が野生株と比べて弱くなるのだ.細胞表面に発現している受容体の総数は変わらないが,その一方で,各一次繊毛上の受容体濃度は薄くなることが観察された.このことから,一次繊毛は受容体やイオンチャネルなどのセンサー分子を集積することで,外部刺激に「敏感」な構造体を達成している,と考えられた.
一次繊毛が重要な生理機能を有することや,ほぼすべての組織に発現していることから想像に難くないことではあるが,一次繊毛の異常はさまざまな疾病 (繊毛病とよぶ)につながることが知られている(概念図).簡単に列挙するだけでも,内臓逆位,嚢胞性腎疾患,網膜変性症,認知障害,多指症,肥満があげられる.特に一次繊毛の異常により発症する多発性嚢胞腎は,500人に1人が罹患する世界で最も多い遺伝病の一つにもかかわらず,治療法がいまだ確立されていない病気で,基礎研究・臨床研究が活発に行われている.
3カルシウム論争
100年以上も前にジマーマンが予見した腎臓における一次繊毛の知覚器官としての実験証明が,2001年と2003年に報告されている7)8).腎臓の培養細胞において,流れ刺激が一次繊毛を曲げ,その後細胞質でカルシウムイオン ( Ca2+)濃度が上昇すること,一方で一次繊毛を化学処理によって摘みとるとこの反応が消失することが観察された.また同2003年,腎臓の細胞で一次繊毛に局在する膜タンパク質PC1やカチオンチャネルPC2(TRPP2ともいう)の変異体を発現したり,機能抑制抗体を添加することで,流れ負荷による細胞内 Ca2+濃度の上昇が抑制されることが報告された9).一連の知見から,一次繊毛にPC1やPC2のようなメカノセンサーを集積することで,機械刺激を受容し,カルシウムの流入が起こるというモデルが提唱されるに至る.そして2012年になると,胚発生段階におけるノードとよばれる領域において,クラウン細胞の一次繊毛が流れ刺激に応答することが体の左右軸の決定に必須であるという報告がなされる10).
ジマーマンの先見の明にはただ驚かされるばかりだが,一見これに反論する論文が2016年にNature誌に発表された11).「一次繊毛はカルシウム応答性のメカノセンサーではない」と題するその論文では,一次繊毛特異的に蓄積するカルシウム応答性の蛍光タンパク質を発現したマウスを用いた実験で,さまざまな強度の機械的な刺激に対して,一次繊毛内でカルシウム上昇が観察されない,もしくは観察される場合であってもそれは細胞質で上昇した Ca2+が一次繊毛に拡散して流れ込んだものであるという報告だ.Nature誌にネガティブな結果が掲載されたこともあり,センセーショナルな論文となった.論文の主旨は,一次繊毛はメカノセンサーかもしれないが,「その初発の情報伝達物質であると思われてきた Ca2+は一次繊毛膜のチャネルを介して流入するのではない」というものであるが,論文が「一次繊毛がメカノセンサーではないと主張している」と多くの人が勘違いしたことも混乱を増強した.筆者の研究室では2015年に流れ刺激依存的な Ca2+濃度の上昇を一次繊毛内特異的に検出したことを報告したが, Ca2+の起源については性急とし議論しなかった12).こうしたいわゆる「メカノセンサー論争」を建設的に議論するため,筆者は2016年にとある生命系国際学会にてセッションをオーガナイズし,関連した研究者が一同に介する機会を設けた.セッション後は演者らとバーに行き,開放的な雰囲気のなかでさらに議論を続けた.そこから生まれた一つの結論は,実験科学に内在する根源的な限界に関する,振り返ってみればある意味当然のものであった.つまり,こうした実験条件ではこういう結果がでるというものである.ただし実験条件は無数に存在するので,ほかの実験条件で同じ結果が得られるという保証はない.それを踏まえて,試行する実験条件ができるだけ一般性・信頼性があるように注意深くデザインする必要があるということである.一次繊毛は特に小さく直接見ることができない現象が多いので,特に細心の注意が必要とされる.
さらに本特集号でこの問題をフォローアップする目的で,前述の論文の筆者らにコメントを求めたので,それを紹介することで読者の思索の助としたい.また横山の稿,水野の稿もぜひ参考にしてほしい.(順番は論文の年代順,敬称略,原文まま,掲載の許可受諾すみ)
● ヘレ・プレトーリアス(Arhus University)
Thus in all fairness, we must consider that the degree of Ca2+ chelation in the cilium is massively altered by introducing substantial Ca2+ binding sites into the narrow space of the primary cilium and that this perturbation may prevent detection of minor influx from the exterior. In this context, one has to consider the Ca2+ binding properties and kinetics of the introduced probes, since not all are suitable to detect subtle changes in the Ca2+ concentration.
● スリア・ナウリ(Chapman University)
With regards to the role of calcium in mechanosensory cilia function, it is important to note that calcium may not act as a sole regulator of signaling proteins localized within the ciliary compartment. However, calcium should be looked at as a part of larger signaling complex that helps regulate signaling pathway.
● マーカス・デリング(University of California, San Francisco)
So where do these findings leave us? First of all we cannot rule out that primary cilia may still function as mechanosensors by employing Ca2+ independent signaling cascades such the YAP/TAZ pathway. However, this type of mechanosensation is not able to explain the physiological function of ciliary ion channels such as polycystins or the underlying molecular mechanism of polycystic kidney diseases. Ultimately the molecular mechanism flow sensation by ciliary ion channels in the primary cilium may be a chemosensation of a yet unknown ligand transported by the flow instead of a direct mechanical activation.
● 濱田博司(理化学研究所)
体の左右を決めるノード流を感知するのは,ノード脇にある動かない一次繊毛です.この一次繊毛が,ケモセンサー or メカノセンサーとして働いているのかは長年の疑問ですが,まだ明確な答えがありません.ノードの不動繊毛を含めて,一次繊毛が Ca2+を介するメカノセンサーではないという論文11)がありますが,われわれはいくつかの理由からメカノセンサーである可能性があると考え,それを検証しています.ただ,この問題に明確な答えを出すためには,なるべく
in vivo に近い実験系を用いる必要がありますから,簡単ではありません.現時点では,一次繊毛がメカノセンサーかどうかは,open questionと考えてよいと思います.
まとめ
一次繊毛に関する知見は蓄積されてきたが,カルシウム論争のような混乱も生じている.その最たる理由が実験の難しさであろう.前述のように一次繊毛は光の回折限界程度の大きさで,光学的な手法の適用が容易ではない.容量にして1 fL以下の一次繊毛においては,1 μMに相当する分子数はたった600分子となる.そして図2からも顕著なように,一次繊毛は細胞質の容量に比べて非常に小さい(1万分の1以下)ため,一次繊毛を精製して生化学的な手法を適用するのも容易ではない.そのため一次繊毛の研究は遺伝学や薬理学的手法に依存せざるを得なかった経緯があるが,それらの実験結果の解釈は一筋縄ではいかない.なぜなら対象となる分子が一次繊毛だけで選択的に機能していればよいが,多くの場合は同分子が一次繊毛外にも発現が確認されていて,対象分子の一次繊毛での役割を純粋に評価することが難しいからである13).そのため,一次繊毛の実験には工夫が必要となるが,こうした技術革新を各論に続く「フォーラム」にまとめたので参照していただきたい.電子顕微鏡の応用やサンプル調製の工夫のしかた(篠原の稿)にはじまり,小さい構造体を詳細に観察する光学的手法(千葉の稿)や,一次繊毛内のタンパク質を網羅的に解析するプロテオミクス技術(石川の稿および池上の稿),そして光ピンセットなどの物理的な技術(加藤の稿)である.これらの先駆的技術およびその組合わせにより,さまざまな分野で一次繊毛の観察や摂動が可能となってきている.今後もこうした異分野との技術開発とも相まって,一次繊毛の研究は加速していくと考えられる.その功績として,一次繊毛の分子に特異的に作用するような薬が創出され,多発性嚢胞腎をはじめとする繊毛症全般を治癒できる日がいつか来るかもしれない.メカニカルセンサー論争の行方も含めて,向こう1世紀にわたって,一次繊毛は研究者の興味を引くことであろう.
※本ウエブサイト上では図2を割愛しています
文献
- Zimmerman KW:Anatomie, 52:552-706, 1898
- Bloodgood RA:Methods Cell Biol, 94:3-52, 2009
- Satir P:Curr Biol, 17:R963-R965, 2007
- Sorokin SP:J Cell Sci, 3:207-230, 1968
- Prosser SL & Morrison CG:J Cell Biol, 208:693-701, 2015
- Mahjoub MR & Stearns T:Curr Biol, 22:1628-1634, 2012
- Praetorius HA & Spring KR:J Membr Biol, 184:71-79, 2001
- Praetorius HA & Spring KR:J Membr Biol, 191:69-76, 2003
- Nauli SM, et al:Nat Genet, 33:129-137, 2003
- Yoshiba S, et al:Science, 338:226-231, 2012
- Delling M, et al:Nature, 531:656-660, 2016
- Su S, et al:Nat Methods, 10:1105-1107, 2013
- Vertii A, et al:EMBO Rep, 16:1275-1287, 2015
- Geerts WJ, et al:J Struct Biol, 176:350-359, 2011
著者プロフィール
井上尊生:東京大学薬学部にてケミカル―バイオロジー,同医学部にてセルバイオロジーを学ぶ.博士号取得後,米スタンフォード大学でシステムズバイオロジーを学び,2008年からジョンズホプキンス大学医学部にて独立.生きた細胞内で活躍する分子プローブを設計,作製および適用し,生物の根源的な謎解きに挑戦する.究極の目標は,繊毛関連疾患の病因に関する新しい洞察を得て,斬新な治療法を開発することである.
 なぜ小さな繊毛を研究するのか
なぜ小さな繊毛を研究するのか







