概論
がん免疫が開くヒト免疫学新時代〜
A new era of human immunology opened by cancer immunology
西川博嘉
Hiroyoshi Nishikawa:Department of Immunology, Nagoya University Graduate School of Medicine/ Division of Cancer Immunology, Research Institute / Exploratory Oncology Research & Clinical Trial Center(EPOC), National Cancer Center(名古屋大学大学院医学系研究科微生物・免疫学講座分子細胞免疫学/国立がん研究センター研究所腫瘍免疫研究分野/先端医療開発センター免疫TR分野)
近年,外科的切除・化学療法・放射線療法といったがん自体を標的とする治療と異なり,宿主の免疫系を活性化することでがん細胞を攻撃,駆逐するがん免疫療法が注目を集めている.とりわけ免疫チェックポイント阻害剤とよばれるCTLA-4やPD-1といった免疫共抑制分子(免疫チェックポイント分子) に対するブロッキング抗体が多くのがん種に対して国内外で臨床応用が進んでいる.免疫チェックポイント阻害剤の臨床応用により,がんがさまざまな免疫抑制機構により免疫系からの攻撃を逃避していることが明らかになってきた.ここでは,本特集でとり上げるがんがもつ免疫抑制機構の研究および今後の臨床展開について概略を述べる.
はじめに
約半数の日本人が悪性腫瘍(がん)に罹患する.がんは日本人の死亡原因の30%を超え年々増加傾向にあり,まさに「国民病」とも言える疾患となっている.がんの治療法として外科的切除,化学療法,放射線療法があり,おのおのの治療分野でめざましい進歩がみられるが,依然としてがんの治癒は困難で,新規治療法の開発が喫緊の課題である.近年,がん免疫療法が第4のがん治療法として注目を集めている.従来のがん治療ががん細胞自体を標的とするのと異なり,がん免疫療法は宿主の免疫系を活性化し,がん細胞を攻撃・駆逐する.前立腺がんに樹状細胞療法を応用したSipuleucel-Tがアメリカ食品医薬品局(FDA)から承認されたのを皮切りに,がん免疫療法は新たな時代に入った.とりわけCTLA-4やPD-1といった免疫共抑制分子(免疫チェックポイント分子)によるT細胞応答の抑制(ブレーキ)をはずすブロッキング抗体(免疫チェックポイント阻害剤)の抗腫瘍効果が証明され,悪性黒色腫,非小細胞肺がん,腎細胞がん,膀胱がん,頭頸部がん,胃がんやホジキンリンパ腫をはじめ,多くのがん種に対して臨床応用が進んでいる.免疫チェックポイント阻害剤の臨床応用により,がん免疫療法の「光と陰」,つまり従来の抗がん剤と比較して進行がん患者でも劇的な効果を示す一方,治療効果の個人差,耐性の出現,副作用などの課題も明らかになってきた.がん免疫療法は依然として発展途上で,がんがどのように免疫系を抑制し生き延びるのか,そのメカニズムを知ることが,がん免疫療法の成功,さらにはがん征圧のための重要な鍵となる.ここでは,がんによる免疫抑制機構に焦点を当て,その解明による新たな治療戦略の現状について概略を述べる.
1がん免疫研究の歴史
免疫系は自己と非自己を識別し,自己には反応せず非自己に反応して排除する.がん細胞は遺伝子変異の蓄積により形成されることから,免疫系はこれらの遺伝子変異に由来するタンパク質を異物(抗原)と認識し,生体内に生じたがん細胞を破壊してがんの進展を抑制していることが現在では明らかになっているものの,「免疫系が生体内に生じたがん細胞を異物として認識し攻撃するか」については長年にわたり議論がなされてきた1).
Coleyが「悪性腫瘍患者で,細菌感染により腫瘍が退縮する」ことがあるのを見出したことが,炎症・免疫をがん治療に応用する試みのはじまりである2).これらの事象をもとに20世紀初頭Ehrlichは免疫系が体内で絶え間なく出現する異常細胞(がん細胞)を排除しないなら,がんの発生は驚くべき頻度となると考え,免疫系ががんから生体を防御していると言う概念を提唱した3).この考えはBurnetとThomasに引き継がれ,1960年代に「生体内では頻繁に細胞に遺伝子異常が引き起こされ悪性細胞が出現するが,これらの危険な悪性細胞は免疫系により認識され排除される」というがん免疫監視機構(cancer immunosurveillance)としてまとめられた4).一方で,Medawarらによる自己免疫寛容機構の解明により,“自己もどき”である腫瘍は免疫系に排除されないという,がんに対する免疫応答に否定的な見解も示された5).1970年代,野生型マウスと比較して胸腺を欠損したヌードマウス(理論上T細胞が存在しないが,実際はT細胞の残存,NK細胞の強い活性化が認められた)で化学発がんに差が認められなかったことが,Stutmanらにより報告される6)と共にColey’s toxinによる治療成果も十分でなかったことから,がんに対する免疫応答の存在が疑問視されるとともに,がん免疫研究の歴史は一時後退した.
その後,遺伝子改変技術などの実験技術の進歩により,種々の免疫関連遺伝子変異動物を用いて発がんへの影響が検討された結果,リンパ球が存在しないRAG(recombination-activating genes)欠損マウスやIFN(interferon)-γやパーフォリンといった抗腫瘍免疫応答にかかわる分子が欠損したマウスでは発がんが促進することが示され,がん免疫監視機構の存在が動物モデルで証明された.さらに,1991年にはヒトがん抗原がBoonらによって同定され7),ヒトにおいてもがん免疫応答の存在が分子的に解明される端緒となった.現在,免疫系が発がんからがんの進展にかかわる過程は,SchreiberやOldらによって「がん免疫編集(cancer immunoediting)」としてまとめられている8)9).
2がん免疫編集とがん免疫療法
(1) がんに対する免疫応答と発がん
免疫系が発がんからがんの進展にかかわる過程は,排除相,平衡相,逃避相の3つに分けられて「がん免疫編集」としてまとめられており8)9),これを理解することはがんに対する免疫療法を理解するうえできわめて重要である.
紫外線や放射線などの外界からのさまざまな刺激により生体内に出現した異常細胞(がん細胞)は細胞の自己修復能により修復されるが,修復できない場合は免疫系から攻撃を受けて排除され,生体の恒常性が維持される(がん免疫監視 = 排除相).しかし,がん細胞は免疫系が存在する環境において生存するのに適した免疫原性が低い(免疫応答が容易に誘導されるがん抗原を有しない)がん細胞をダーウィンの自然選択説的に選択することで,免疫系からの攻撃を回避して生体内に生存可能となるが,この状態では無限に増殖できない(平衡相).そこで,本来は不適切,もしくは過剰な免疫応答を抑制することで生体の恒常性を維持するのに重要な働きをしている免疫抑制機構をがん組織に取り込み,積極的に抗腫瘍免疫応答を抑制する環境をつくり上げて免疫系からの攻撃を逃避することで無限に増殖し(逃避相),臨床的「がん」となる.よって,臨床的に診断される「がん」は,免疫学的に選択され,多様な免疫逃避機構を確立した“編集(edit)された”がん細胞の集団となっている8)9).つまり,がん免疫療法を成功させるには,がん細胞にすでに構築された免疫抑制ネットワークを解除するとともに免疫原性の低いがん細胞に対して強力な免疫応答を誘導する必要がある.
(2) がん免疫編集から見たcold tumorとhot tumor
免疫応答をがん治療に応用する試みは,BCG(Bacille de Calmette et Guérin)の膀胱がん治療やIL-2などのサイトカイン療法の悪性黒色腫,腎細胞がんなどへの応用例があるものの,長年にわたりがん治療の第4の柱としてのがん免疫療法の成功には結びつかなかった.さらにヒトがん抗原の同定により,がん抗原分子を投与することで抗腫瘍免疫応答を賦活化する試み(がんワクチン療法)がなされたが,期待されたような臨床効果は認められなかった.このとき用いられたがん抗原は,多くのがん患者で共通して発現が認められる自己由来抗原分子(shared antigen)で,自己由来のこれらの抗原に対しては,胸腺での正および負の選択により自己免疫寛容が成立している5)10).さらに近年の抗原特異的T細胞の解析技術の進歩により,胸腺での負の選択を免れた自己反応性T細胞が,末梢で制御性T細胞に抑制され,不可逆な不応答状態に陥ることで免疫寛容が維持されていることも示されている11).よって自己由来抗原分子(shared antigen)に対するCD8+T細胞を活性化して有効な抗腫瘍免疫応答を誘導するには,制御性T細胞を除去したうえでCD8+T細胞を活性化に導くなどの複合免疫療法が必須である.
一方,がん細胞に蓄積する遺伝子変異に由来するタンパク質は,生体にとって新たに出現した異物(抗原)であり,がん特異抗原(tumor-specific antigens, TSA=neoantigen:ネオ抗原)とよばれる.これらの抗原は,免疫系とりわけCD8+T細胞の標的となるため,ネオ抗原が多数存在するがんではがん局所にCD8+T細胞が多数存在すると考えられる10).しかし,がん細胞は,発がんの過程でCD8+T細胞からの攻撃を逃避する必要があることから免疫抑制細胞および免疫抑制関連分子発現も同時に認められるimmunogenic tumor(hot tumor)になると考えられる12)13).さらに,がん免疫編集説に従えば,これらの異物と認識される免疫原性の高い抗原分子は,本来は発がん過程で排除されているべき抗原と考えられる.よって平衡相において,これらの免疫原性が高いがん抗原が脱落しなかったがん細胞は抗原提示機構やIFN-γなどのエフェクター分子の受容体などに異常があり,免疫原性が高いがん抗原が抗原提示されないか,それらのがん抗原に対する免疫系の攻撃に対して不応答性のがん細胞が選択されてnon-immunogenic tumor(cold tumor)になっていることも考えられる14)15).
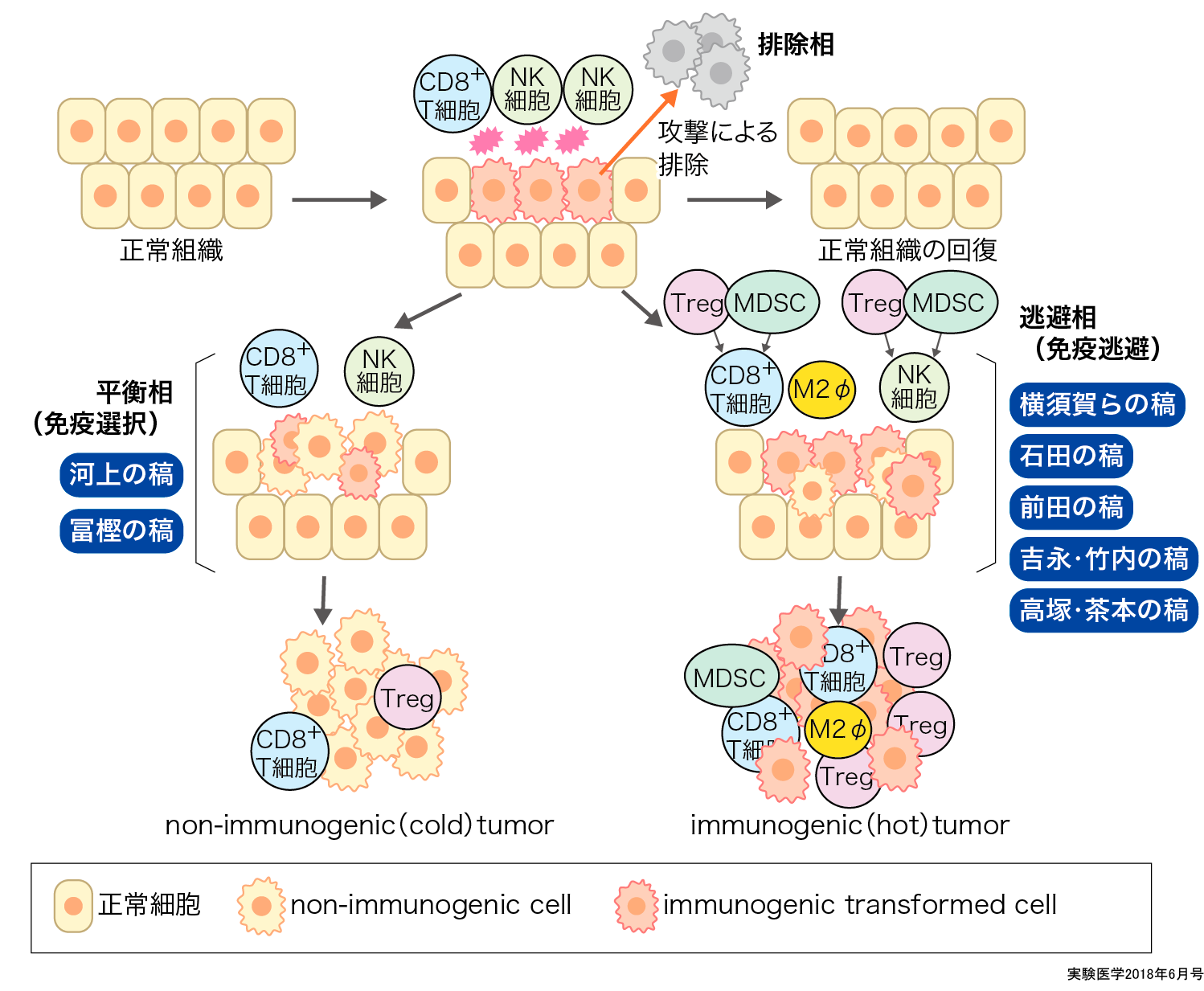
発がんと免疫系とのかかわりは排除相,平衡相,逃避相の3つに分けられている.がん免疫編集での平衡相,逃避相にかかわる免疫選択(primary resistance)と免疫逃避(adaptive resistance)はオーバーラップして作動し,発がんに至る.
すなわち,
① がん細胞は,自身が免疫系の排除から免れるため免疫原性の高い抗原を脱落させ,“自己もどき”となることで免疫系から逃避して増殖する(primary resistance)(河上の稿,冨樫の稿参照)
② がん細胞は,制御性T細胞(Treg),骨髄由来免疫抑制細胞(MDSC)や免疫チェックポイント分子などによる免疫抑制を積極的に活用し抗腫瘍免疫応答を抑制する(adaptive resistance)(横須賀らの稿,石田の稿,前田の稿,吉永・竹内の稿,高塚・茶本の稿参照)
という2つの機構は,マウスモデルで示されたように① primary resistanceが作動し平衡相に達した後に② adaptive resistanceが作動するというような段階的な進行というよりも,ヒトがんでは両者がオーバーラップして作動していると考えられる(概念図).すなわち,個々のがん患者の臨床的「がん」は,それぞれの機構への依存性が異なり,① primary resistanceの機構が主に発がんにかかわっているnon-immunogenic tumor(cold tumor)と,② adaptive resistanceが主なimmunogenic tumor(hot tumor)が存在する15).②が主ながんはimmunogenic tumor(hot tumor)で,ネオ抗原が多数存在しており,抗腫瘍免疫応答はready to goの状態であり,免疫チェックポイント分子などによる免疫抑制を外すことで十分な抗腫瘍免疫応答を活性化できると考えられる.①が主ながんでは,現状のがん免疫療法の対象となりにくく,前述の自己由来抗原分子(shared antigen)に対するCD8+T細胞に対する免疫応答を誘導するなどの検討が必要となる.
② adaptive resistanceが主ながんでの一例が,CD8+T細胞などのエフェクターT細胞が浸潤してIFN-γを分泌したことによりがん局所に誘導されるPD-1リガンド(PD-L1)発現である.つまり,エフェクターT細胞の攻撃を受けたため,このままでは発がんにいたることができないことから,PD-L1発現を誘導してエフェクターT細胞からの攻撃を免れて発がんしてきた可能性が報告されている12).事実,ヒト悪性腫瘍の網羅的遺伝子解析データでも,細胞傷害関連分子は免疫抑制分子とともにがん組織に発現していることが示されているように,がん局所ではがんを攻撃し腫瘍増殖抑制に働く細胞・分子は,これらの抗腫瘍免疫応答を抑制して腫瘍増殖を助ける免疫抑制細胞・分子と混在して存在し,immunogenic tumor(hot tumor)となっていることが示されている(図)16).
(3) がん細胞の直接的な免疫系への作用によるcold tumor
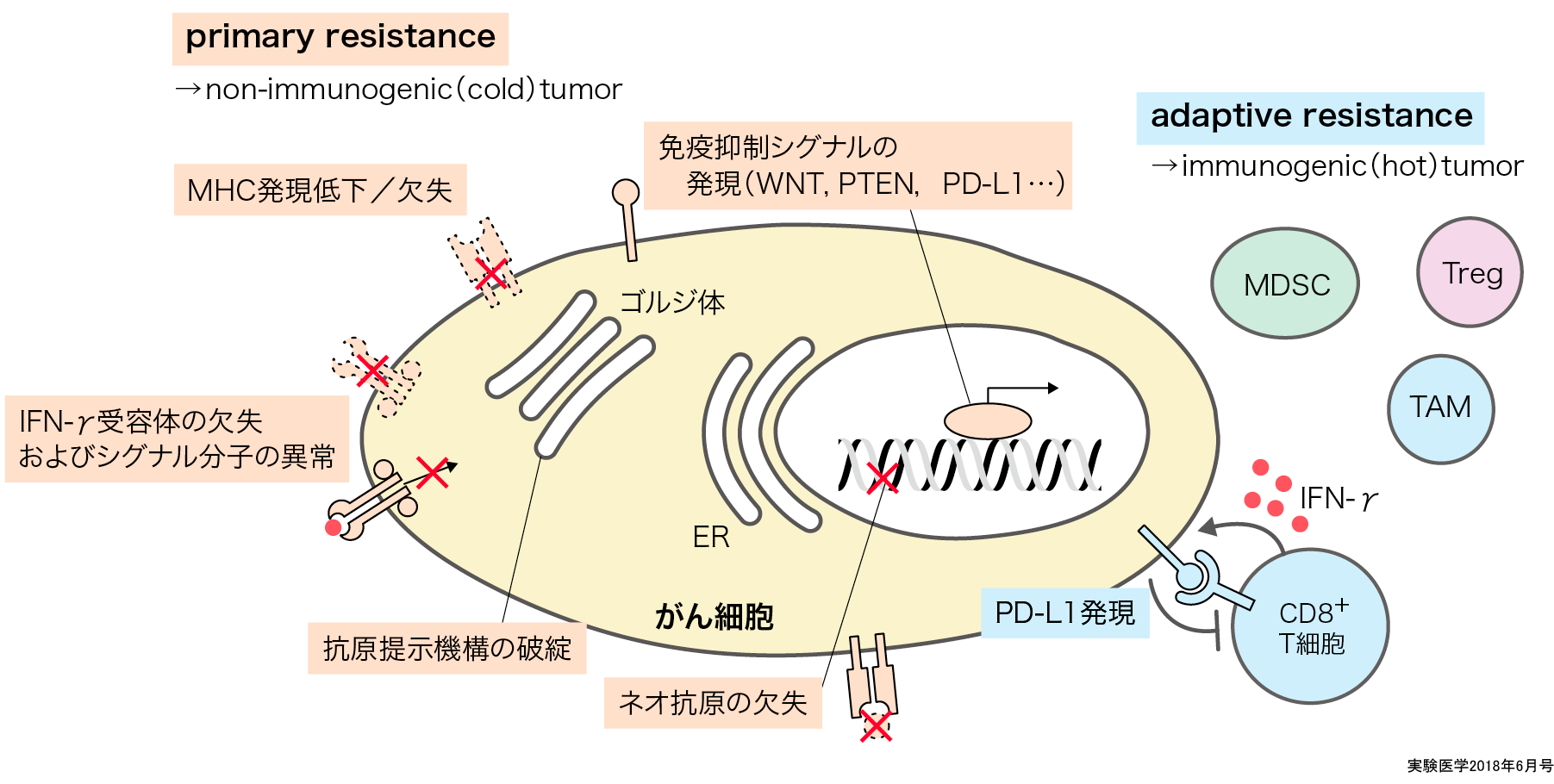
primary resistance(赤)が強く働いた腫瘍は主にnon-immunogenic tumor(cold tumor)となり,adaptive resistance(青)が主に働いた腫瘍はimmunogenic tumor(hot tumor)となる.
発がん過程で生じた遺伝子異常に伴うシグナル(ドライバー変異など)によりがん細胞自身が抗腫瘍免疫応答を抑制していることが明らかになっている.つまり,平衡相では免疫原性の高い抗原分子などを脱落したがん細胞が選択されることに加えて,直接的に抗腫瘍免疫応答を抑制できる遺伝子異常をもったがん細胞が選択されている可能性が示唆される.WNT/β-
catenin17),PTEN18),MYC19)やFAK(focal adhision kinase)20)分子のがん細胞での活性化が,CD8+T細胞の浸潤抑制,免疫チェックポイント分子の高発現やTreg浸潤促進に関連することが示されている.特にWNT/β-catenin活性化は,ATF3を誘導し,それにより
3がん免疫療法により誘導される治療抵抗性
がん免疫療法の臨床応用により,治療抵抗性が獲得されることが明らかになってきた.これは,がん免疫療法の治療効果の特徴である長期の臨床効果(Kaplan-Meier Curveのtail plateau)を阻害するものであり,十分な注意が必要である.特にネオ抗原の欠失や抗原提示機構およびIFN-γシグナルにかかわる異常ががん免疫療法を施行中に獲得されることが報告されている(acquired resistance)15).これらの異常は免疫系の攻撃から免れるためにがん細胞が発がんの過程で獲得する機構(primary resistance)でもあることから,がん免疫療法によりこれらのがん細胞が新たに出現したのか,すでに存在していたがん細胞が選択されたのかは今後の検討課題である.
おわりに
がん免疫療法が臨床応用されるにつれ,がん組織のゲノム解析,免疫応答解析のみならずゲノム不均一性の解析や代謝解析まで幅広い解析の必要性が示唆されている.がん細胞の遺伝子不安定性は,がん抗原を提供する一方で,がん免疫療法に対する新たな治療抵抗性にかかわることが示されてきている.免疫チェックポイント阻害剤に抵抗性となった腫瘍ではHLA classⅠやIFN-γシグナルに関連する遺伝子変異もしくは欠損が存在することやCD8+T細胞に認識されていた遺伝子変異に由来するネオ抗原の発現が低下することも示された.本特集ではこれらのがんがもつ抵抗性について,とりわけ免疫抑制機構に焦点を当ててとり上げる.臨床効果につながる抗腫瘍免疫応答の本態およびそれらに対する免疫抑制機構の解明には,基礎,臨床を超えたさまざまな研究分野の横断的な検討が必要である.
文献
- Pardoll D:Annu Rev Immunol, 21:807-839, 2003
- Coley WB:Ann Surg, 14:199-220, 1891
- Ehrlich P:Ned Tijdschr Geneeskd, 5:273–290, 1909
- Burnet FM:Lancet, 1:1171-1174, 1967
- Billingham RE, et al:Nature, 172:603-606, 1953
- Stutman O:J Natl Cancer Inst, 62:353-358, 1979
- van der Bruggen P, et al:Science, 254:1643-1647, 1991
- Dunn GP, et al:Nat Immunol, 3:991-998, 2002
- Schreiber RD, et al:Science, 331:1565-1570, 2011
- Gubin MM, et al:J Clin Invest, 125:3413-3421, 2015
- Maeda Y, et al:Science, 346:1536-1540, 2014
- Spranger S, et al:Sci Transl Med, 5:200ra116, 2013
- Rooney MS, et al:Cell, 160:48-61, 2015
- Zaretsky JM, et al:N Engl J Med, 375:819-829, 2016
- Sharma P, et al:Cell, 168:707-723, 2017
- Hugo W, et al:Cell, 165:35-44, 2016
- Spranger S, et al:Nature, 523:231-235, 2015
- Peng W, et al:Cancer Discov, 6:202-216, 2016
- Casey SC, et al:Science, 352:227-231, 2016
- Jiang H, et al:Nat Med, 22:851-860, 2016
著者プロフィール
西川博嘉:1995年三重大学医学部卒業.三重大学医学部附属病院等にて臨床研修の後,’98年三重大学大学院医学系研究科博士課程内科学専攻入学.2002年同修了(医学博士).’03年Memorial Sloan Kettering Cancer Center リサーチフェロー,’06年三重大学大学院医学系研究科病態解明医学講座 講師,’10年大阪大学免疫学フロンティア研究センター実験免疫学 特任准教授,’12年Roswell Park Cancer Institute Adjunct Associate Professor(兼任)を経て,’15年より国立がん研究センター研究所 腫瘍免疫研究分野/先端医療開発センター免疫トランスレーショナルリサーチ(TR)分野 分野長,’16年より名古屋大学大学院医学系研究科微生物・免疫学講座分子細胞免疫学 教授とをクロスアポイントメントで免疫学,とりわけ腫瘍免疫学の基礎研究からTRまでを進めている.近年はがん局所における免疫抑制ネットワークの本態解明とそれらを標的とした新規がん免疫療法の開発を行っている.
発がんと免疫系とのかかわりは排除相,平衡相,逃避相の3つに分けられている.がん免疫編集での平衡相,逃避相にかかわる免疫選択(primary resistance)と免疫逃避(adaptive resistance)はオーバーラップして作動し,発がんに至る.
primary resistance(赤)が強く働いた腫瘍は主にnon-immunogenic tumor(cold tumor)となり,adaptive resistance(青)が主に働いた腫瘍はimmunogenic tumor(hot tumor)となる.










