
概論
T細胞機能不全をイムノメタボリズムから理解する
T cell dysfunction and immunometabolism
山下政克
Masakatsu Yamashita:Department of Immunology, Graduate School of Medicine, Ehime University(愛媛大学大学院医学系研究科免疫学講座)
近年,T細胞の代表的な機能不全である,疲弊,不応答(アナジー)や細胞老化を制御する転写 調節因子が同定され,T細胞の機能不全の分子メカニズムが解明されつつある.一方で,イムノメタボリズムを介した制御がT細胞の分化や機能において重要であることが多くの研究から明らかとなっている.さらに,イムノメタボリズムの不具合が,T細胞機能不全とそれによるがん免疫応答や感染免疫応答の低下に深くかかわっていることもわかってきた.本特集では,T細胞疲弊と老化の誘導メカニズムの最新の知見について,イムノメタボリズムと転写因子の関与を中心に紹介する.
はじめに
がん免疫応答の低下や慢性感染症とイムノメタボリズム,T細胞の機能不全は,密接に関係している.代表的なT細胞機能不全として,疲弊,アナジーと老化がある.現在のところ,疲弊と不応答(アナジー)は,その誘導条件には後述するような違いはあるものの,非常によく似た解除可能な機能不全であり,解除により腫瘍免疫や感染免疫応答の賦活化が期待される.これに対し,老化T細胞は不可逆的な機能不全であり,老化随伴分泌現象(senescence-associated secretary phenotype,SASP)により炎症性サイトカインを高産生し,自己免疫疾患に代表される慢性炎症疾患の病態形成に関与している可能性が示されている.細胞内の代謝状態については,疲弊やアナジーでは解糖能の低下が認められるのに対し,老化細胞は高い代謝状態が保たれている.本稿では,T細胞の疲弊,アナジー,老化の特徴と誘導機構について,イムノメタボリズムと転写因子の観点から概説する.
1T細胞イムノメタボリズム
❶ T細胞活性化と代謝リプログラミング
細胞内代謝は,比較的安定的で変化に乏しいと考えられてきた.しかし,がん細胞を中心とした近年の研究により,細胞の代謝状態は細胞外環境の変化などの影響を受けて劇的に変化し,その適応異常が疾患発症の引き金となることがわかってきた.代謝適応に伴う代謝経路の網羅的かつ協調的な変化は,代謝リプログラミングとよばれ,細胞の分化や環境変化に適応 するための細胞機能の変化に必須であると考えられている.
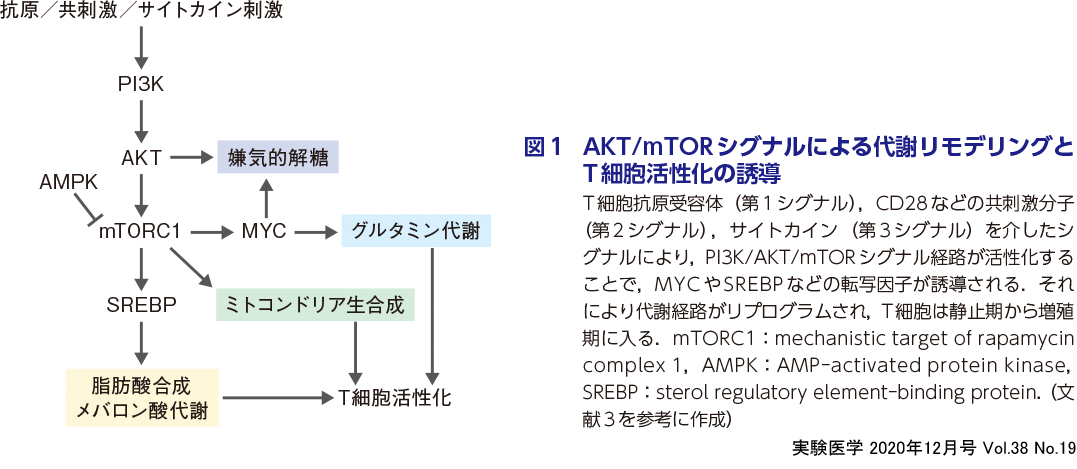
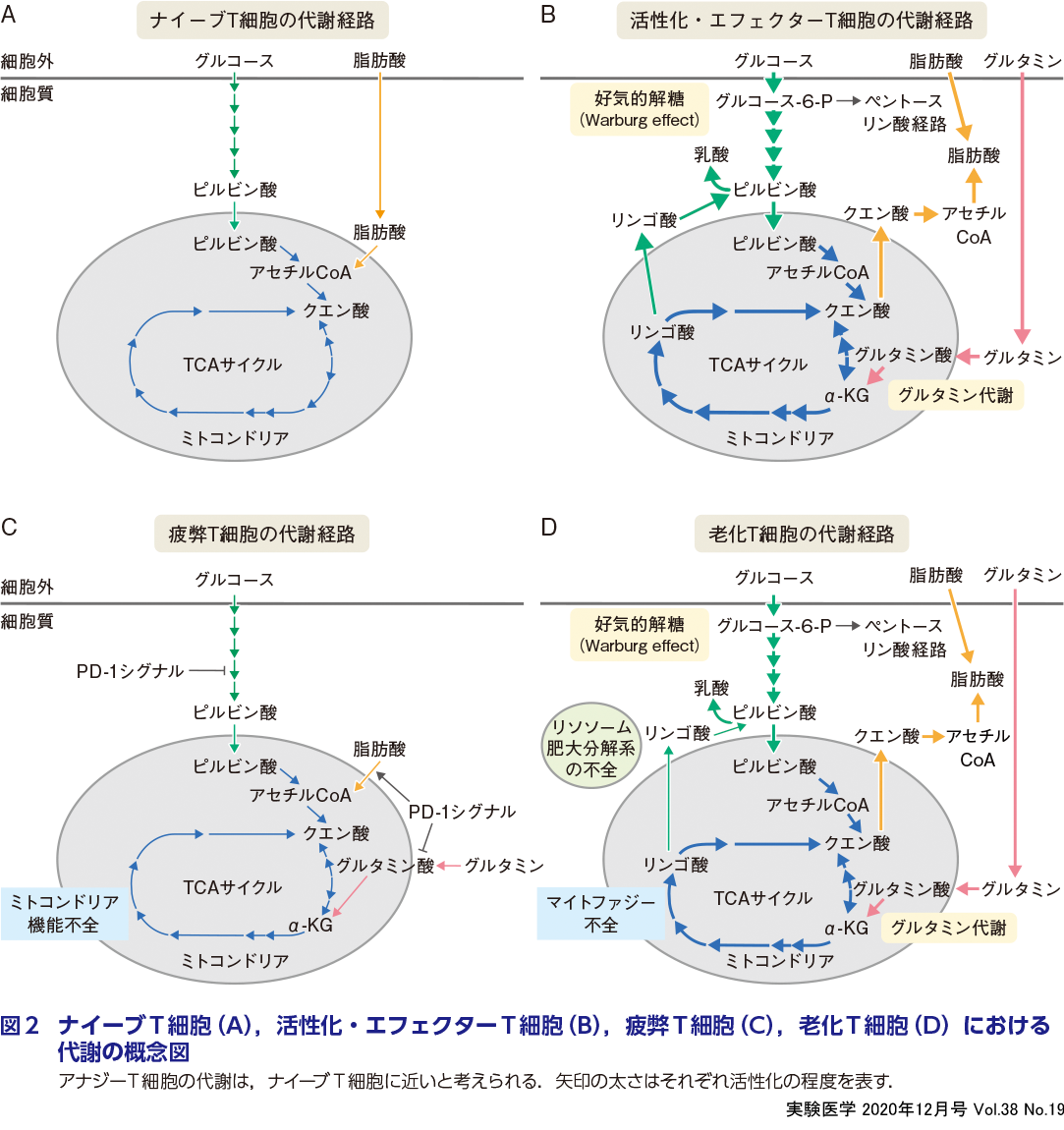
T細胞では,抗原認識(第1シグナル),共刺激(第2シグナル)やサイトカイン刺激(第3シグナル)によって細胞内エネルギー代謝経路が劇的に変化し,それが分化,機能の変化を 制御することが報告されている1)2)(図1).ナイーブT細胞は,細胞外から少量のグルコースや脂肪酸を取り込み,主にクエン酸(TCA)サイクルを使った酸化的リン酸化によりATPをつくり出して生存を維持している(図2A).一方,活性化T細胞では,グルコーストランスポーターの発現上昇に伴って細胞外からのグルコースの取り込みが大幅に上昇し,活発に増殖するがん細胞と同様にWarburg effect(ワールブルグ効果)が誘導される.すなわち,豊富な酸素が存在し,ミトコンドリアで酸化的リン酸化が十分に行えるにもかかわらず,嫌気的解糖が行われる(図2B).さらに,活性化T細胞では,細胞外からのグルタミンの取り込みも著しく増加し,グルタミン代謝が活発化する(図2B).グルタミン代謝の代謝産物であるα-ケトグルタル酸(α-KG)は,TCAサイクルで使用されるとともに,脂質や核酸などのマクロ分子の合成にも利用される.さらに,活性化T細胞では,脂肪酸合成も活発化するだけでなく, 細胞外からの遊離脂肪酸の取り込みも増加し,T細胞増殖を促進する(図2B).
❷ 活性化T細胞の代謝調節機構
T細胞の代謝リプログラミングでは,T細胞抗原受容体(TCR)下流のシグナル伝達経路が重要な働きを担っている3).抗原刺激によって誘導されるPI3K/AKT/mTORシグナルの活性化は,ナイーブT細胞が静止期から活性化・増殖期に入るために必須であり,解糖系,グルタミン代謝,ミトコンドリア生合成,脂肪酸合成,メバロン酸代謝の活性化を誘導する(図1).PI3K/AKT/mTORシグナルによる代謝リプログラミングは,MYCやSREBPといった転写因子を介して誘導される.また,抗原刺激によりAMPKの活性化も誘導される.T細胞の活性化初期において,AMPKの活性化は,mTORC1シグナルの活性化を制限し,増殖期に必要なグルコースの同化が早期に開始されるのを抑制することでT細胞の活性化を調節していると考えられている.
❸ T細胞代謝リプログラムとエピジェネティクス,シグナル伝達
T細胞の代謝リプログラミングにより,T細胞のエピゲノム状態が大きく変動することが 予想される.これは,代謝産物の多くがエピジェネティック制御酵素の基質や補因子であることに由来する.また,代謝レベルはATPの産生レベル,消費レベルと関連する.ATPは, タンパク質リン酸化酵素やユビキチン化酵素の活性化に必要であり,細胞内シグナル伝達も大きく影響されることが考えられる.
2機能不全T細胞の誘導機構と特徴
❶ T細胞クローナルアナジー
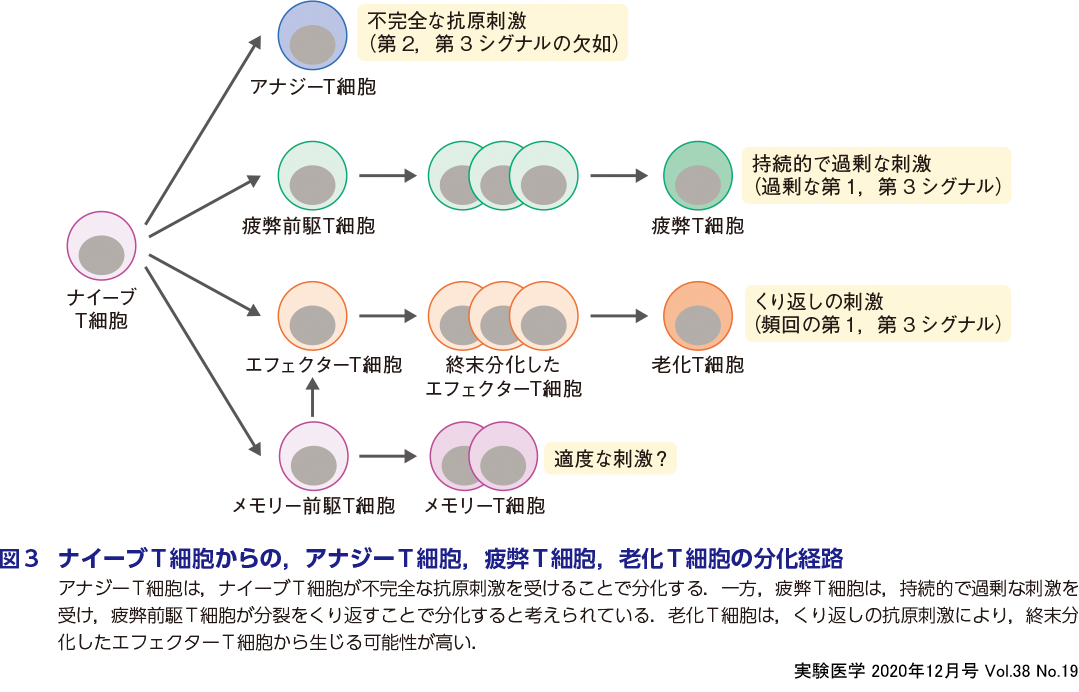
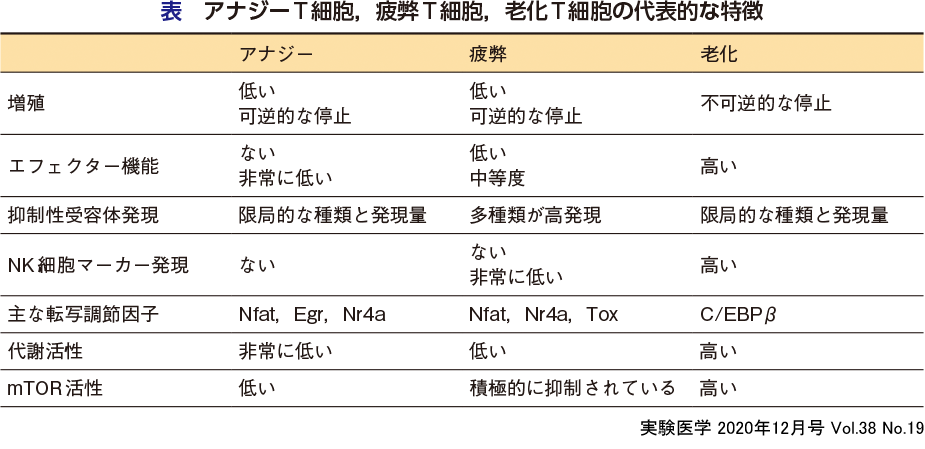
前述したように,T細胞が活性化するためには,TCRを介した第1シグナルと,CD28を 介して伝えられる補助シグナル(第2シグナル)が必要であるが,第2シグナルがない状態で第1シグナルを受けとったT細胞において,2回目の刺激に対して増殖能とIL-2,IFN-γやTNF-αなどのサイトカイン産生の低下が誘導される現象をクローナルアナジー(アナジー,アナージー)とよんでいる4)5)(図3,表).T細胞アナジーは,転写因子AP-1がない状態で,転写因子Nfatがホモダイマーを形成して活性化により誘導されることが報告されている.細胞増殖やサイトカイン産生の低下などのアナジーの特徴は,in vitroにおいて,T細胞の活性化によく使われるPMA(ホルボール-12-ミリスタート-13-アセタート)刺激やIL-2の添加で解除されることから,アナジーは可逆的な現象であると言える.
細胞表面分子や転写調節因子の発現様式から見た場合,アナジーと疲弊は似た現象と考えられるが(安藤らの稿),詳細な比較は行われておらず,今後,アナジーT細胞と疲弊T細胞のエピゲノム状態,メタボローム状態を比較することが必要である.アナジーT細胞表面では,CTLA-4やPD-1の発現が上昇するが,これらの分子は疲弊でも誘導される.また,アナジーT細胞で高発現する転写因子には,Egr2/Egr3,Nr4a1などがあるが,これらの転写因子はいずれもNfatの標的分子であり,疲弊T細胞や制御性T細胞においても発現していることから,T細胞免疫寛容状態の誘導・維持に広く必要な転写因子と捉えることができる.
また,アナジーT細胞では活性化T細胞に比べ,解糖系の著しい低下が認められる.これは,T細胞代謝リプログラミングにCD28を介した第2シグナルが必要なため,アナジー誘導条件では代謝リプログラミングが誘導されないためであると考えられる.
❷ T細胞疲弊
T細胞疲弊は,慢性感染症(玉井・倉知の稿)や腫瘍局所(西田・鵜殿の稿)においてT細胞が持続的に過剰な第1,第3シグナルを受けることで誘導される機能不全であると考えられている5)〜7)(図3,表).疲弊T細胞の特徴は,増殖能やエフェクター機能(IFN-γ,TNF-αやIL-2などのサイトカイン産生や細胞傷害活性)の低下,抑制性受容体(PD-1,Tim-3,LAG-3,TGITなど)の発現増加である.
疲弊T細胞の誘導にかかわる転写因子としては,Nfat,核内受容体であるNr4aファミリー分子(Nr4a1,Nr4a2,Nr4a3),Tcf7やBlimp1などが知られていたが,最近,T細胞疲弊誘導のマスター転写因子としてHMGボックス転写因子であるToxファミリー分子が報告された(安藤らの稿).Nr4aとToxの発現誘導は,いずれもNfatに依存していることから,Nfat依存的にNr4aやToxが誘導されることがT細胞疲弊の原因となっていると考えられている.Nr4a1の誘導やAP-1依存的な転写活性の低下はアナジーT細胞でも認められるが,Toxの関与については明らかになっていない.Nr4aやToxは,AP-1やNF-κB(p65)依存的な転写を抑制することで疲弊状態を維持している.最近,転写抑制因子Bach2の発現が高いことがCD8 T細胞疲弊の誘導に必要であることが示された8).Bach2は,AP-1配列に結合し,AP-1依存的な転写活性化を抑制することが報告されており,Nfat,Nr4aやToxとの関係の解析が待たれる.
アナジーT細胞と同様に疲弊したT細胞では,エフェクターT細胞に比べ解糖能が低下している(西田・鵜殿の稿).疲弊T細胞のマーカーの一つでもある免疫チェックポイント分子PD-1を介したシグナルは,解糖系の活性化を抑制するとともに,脂肪酸酸化の遷延を引き起こし,T細胞応答を負に制御する9)(図2C).PD-1を介した解糖能の抑制は,AKT/mTORシグナルの抑制に依存しており,T細胞疲弊の要因となる(平野・茶本の稿).
❸ T細胞老化
老化は,これまで加齢に伴う胸腺の退縮や長期にわたる頻回な抗原刺激によって細胞分裂がくり返され,T細胞が分裂限界を迎えることで誘導されるとされてきたが(濵﨑の稿),最近の研究により,疲弊と同様に腫瘍微小環境で積極的に誘導されることも報告されている10).アナジーや疲弊とは異なり,老化T細胞は,終末分化したT細胞と形質がよく似ている(図3,表).
老化T細胞での細胞周期の停止は不可逆的なものである.また,老化CD8 T細胞では, IFN-γなどのサイトカイン産生や細胞傷害活性は低下せず,逆にSASPとよばれる炎症性因子,炎症性サイトカイン・ケモカインの高産生が誘導される.SASPもまた,老化をきたした細胞に共通した特徴である.老化T細胞に特徴的な細胞表面分子の発現様式には,CD28やCD27などの共刺激分子の発現消失,活性化型NK受容体とその活性化型アダプター分子,抑制性受容体の発現増加があげられる.
老化T細胞分化の転写・エピゲノム調節は,アナジーや疲弊に比べて解析が遅れている. 老化T細胞において,C/EBPβやGata4の発現増加,NF-κB(p65)の恒常的な活性化, クロマチン構造の維持にかかわるSatb1発現の低下などが報告されている7).また,Bach2の発現低下は,活性化後の急速なエフェクターT細胞の終末分化と細胞老化を誘導する11).疲弊T細胞ではBach2発現維持が必要であり,老化T細胞はBach2の発現低下によって誘導されることは,これら2種類のT細胞機能不全を考えるうえで興味深い.
アナジーや疲弊したT細胞では,AKT/mTORの活性が抑制され,解糖能が低下しているのに対し,老化T細胞ではAKT/mTORシグナルの活性化亢進・遷延による解糖系やグルタミン代謝の持続的な活性化が認められる7)(鈴木らの稿)(図2D).老化細胞における,エネルギー代謝経路の活性化亢進は,SASPなどのエフェクター機能を支えるうえで重要である.さらに,老化した細胞の特徴として,senescence-associated β-galactosidase(SA-βgal)活性の上昇とユビキチン化タンパク質やリポフスチンの蓄積がある5).SA-βgalはリソソーム酵素であり,老化細胞ではリソソームの肥大化と活性上昇が認められる.また,リポフスチンは不飽和脂肪酸の過酸化によりリソソーム内に蓄積される残余物質である.これらのことは,細胞内分解系の障害とT細胞老化が密接に関連していることを示している(渡邊らの稿).
おわりに
近年,T細胞の分化,機能不全に関する転写,エピゲノム研究は著しい進歩を見せている.また,メタボローム解析法の進歩により,T細胞におけるイムノメタボリズム研究も急速に進んだ.しかしながら,これら2つの研究領域を統合する研究はまだそれほど多くはない.また,T細胞疲弊や老化の研究の多くはCD8 T細胞を用いたものであり,CD4 T細胞でも誘導機構や形質はCD8 T細胞と同じなのかなど解明されていない点は多い.
ごく最近,T細胞のミトコンドリア機能を低下させメタボリズムに異常をきたしたマウスでは,個体老化が進み,さまざまな加齢関連疾患が発症することが示された12).また,老化T細胞に発現するCD153を標的としたペプチドワクチンの投与により,糖尿病モデルマウスの病態が改善することも報告されている13).このことは,T細胞代謝を調節し,疲弊や老化を制御することが,がんや慢性感染症だけでなく,さまざまな加齢関連疾患の治療につながることを意味している.しかしその一方で,老化T細胞は高齢者の生体防御に関与している可能性も報告されている14).おもしろいことに,スーパーセンチナリアン(110歳に到達した超長寿者)において,細胞傷害活性を有するCD4 T細胞が増加していることが報告されている15).今後到来する超高齢化社会において,イムノメタボリズムを制御し,T細胞免疫機能を適切に調節するための研究と新規技術の開発が,健康寿命の延伸に重要になると考えられる.
文献
- Geltink RIK, et al:Annu Rev Immunol, 36:461-488, 2018
- Zhang L & Romero P:Trends Mol Med, 24:30-48, 2018
- Chapman NM, et al:Nat Rev Immunol, 20:55-70, 2020
- Zheng Y, et al:EMBO Rep., 9:50-55, 2008
- Huff WX, et al:Int J Mol Sci, 20:doi:10.3390/ijms20112810, 2019
- Wherry EJ & Kurachi M:Nat Rev Immunol, 15:486-499, 2015
- Goronzy JJ & Weyand CM:Nat Rev Immunol, 19:573-583, 2019
- Utzschneider DT, et al:Nat Immunol, 21:1256-1266, 2020
- Delgoffe GM & Powell JD:Mol Immunol, 68:492-496, 2015
- Xia A, et al:Front Immunol, 10:1719, 2019
- Kuwahara M, et al:Nat Commun, 5:3555, 2014
- Desdín-Micó G, et al:Science, 368:1371-1376, 2020
- Yoshida S, et al:Nat Commun, 11:2482, 2020
- Pereira BI, et al:Nat Immunol, 21:684-694, 2020
- Hashimoto K, et al:Proc Natl Acad Sci U S A, 116:24242-24251, 2019
著者プロフィール
山下政克:1989年筑波大学第二学群農林学類卒業.’91年大阪大学医学系研究科医科学修士課程修了.’91年藤沢薬品工業 ㈱ 研究員.’98年に薬学博士取得後,千葉大学医学研究院(寄附講座助手,さきがけ研究21専任研究員,講師,准教授),(公財)かずさDNA研究所ゲノム医学研究所を経て,2012年より現職(愛媛大学大学院医学系研究科 教授).T細胞の代謝-エピゲノムを制御することで疾患治療を行う方法論を確立することをめざし研究を行っている.









