
概論
多様なミクログリアの機能と細胞特性
Diverse microglial functions and their cellular properties
増田隆博
Takahiro Masuda:Department of Molecular and System Pharmacology, Graduate
School of Pharmaceutical Sciences, Kyushu
University(九州大学大学院薬学研究院薬理学分野)
きわめて高度かつ華麗な中枢神経系の機能を正確に維持するために欠かすことのできない細胞がいる.それが,今回の特集で紹介する中枢神経系の主要免疫細胞「ミクログリア」である.本稿では,長年の研究から明らかになってきたミクログリアの細胞特性や機能,他の免疫細胞との関係,中枢神経系疾患への関与や治療標的としての可能性について概説するとともに,本特集の概要について簡単に紹介する.
はじめに
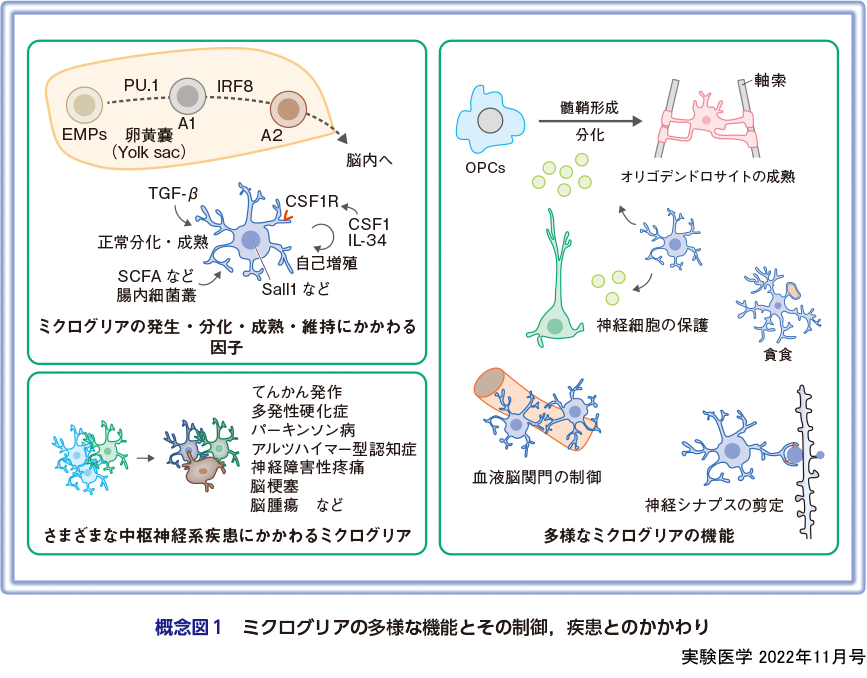
ミクログリアは,脳や脊髄といった中枢神経系組織の実質内に存在する免疫細胞である.アストロサイトやオリゴデンドロサイトとともに,脳内主要グリア細胞として認識されている一方で,本来の機能的分類から脳内マクロファージとしても知られている.今から約100年前,スペイン人神経科学者Pío del Río-Hortega博士は,炭酸銀染色法を用いて中枢神経系を構成する細胞群を発見し,ミクログリアと命名した.ミクログリアの形態や基本的な遺伝子発現プロファイルは,進化学的にみても高度に保存されており1),各動物種で普遍的に重要な機能を担ってきたと考えられる.近年,各種オミクス解析法やイメージング技術の進歩に伴い,ミクログリアの新たな機能的側面が次々に明らかになってきており,死細胞の除去や組織炎症の制御はもちろんのこと,神経シナプスの剪定や,神経新生や髄鞘形成の促進といった正常組織形成過程への関与など,典型的な免疫細胞の機能の域を超え,ほぼすべての脳内生理現象にかかわるとまで考えられるようになった(概念図1)2).
1ミクログリアの発生・維持・老化
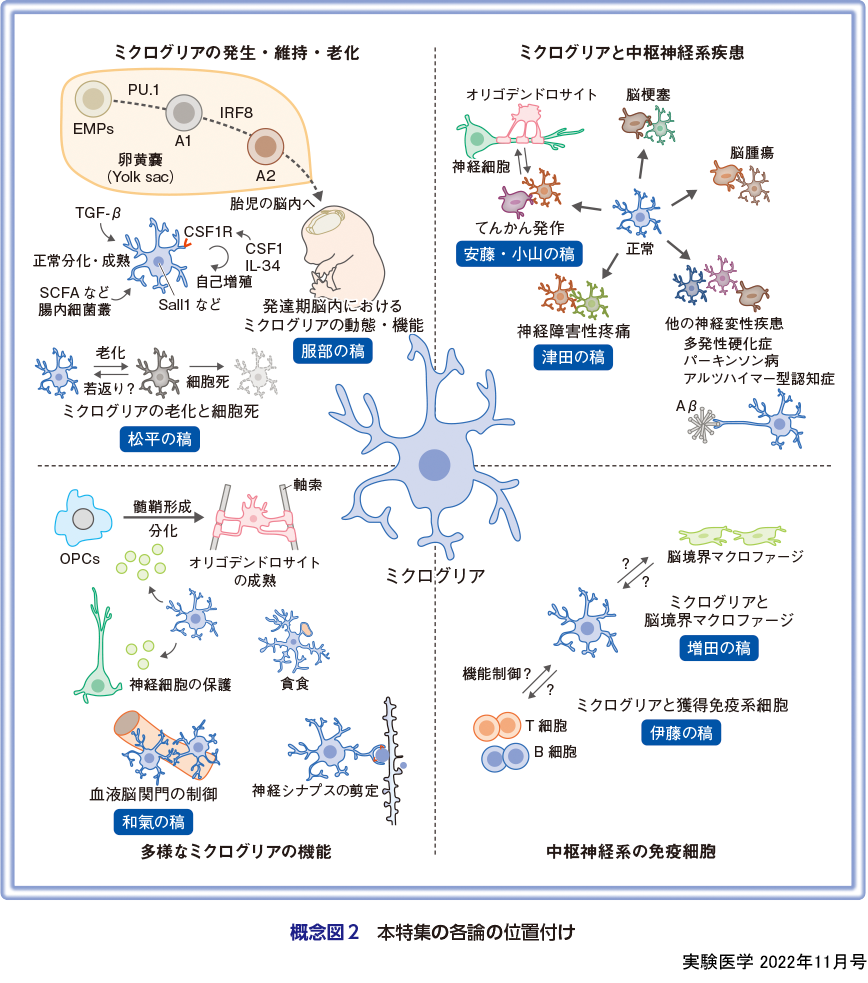
ミクログリアが存在する中枢神経系は,神経細胞のみならず,多種多様な細胞種によって構成された複雑な構造体である.そうした複雑性の構築は,胎生期に始まり,決められたタイミングで多様な細胞が導入され,中枢神経系組織を形成していく.ミクログリアの起源は,胎生早期に卵黄嚢(yolk sac)のblood islandで出現するEMPs(erythro-myeloid progenitors)であると考えられている.EMPsは,転写因子PU.1およびIRF8依存的に,A1細胞,さらにはA2細胞というマクロファージ前駆細胞へと分化した後,脳の形成初期(マウスでは胎生9.5日以降,ヒトでは妊娠4.5週以降)に,胎児の循環系を介して脳周囲に到達し,その後実質内に浸潤し分化成熟する3).発達過程の脳内において,ミクログリアは非常にダイナミックな動態を示し,その時々で重要な役割を果たす.本特集では,脳発達過程におけるミクログリアの動態と機能について概説する(服部の稿).
一方,脳定着後は,少なくとも正常状態では,末梢血から骨髄由来細胞の供給を受けることなく,低頻度の増殖と細胞死をくり返しながら組織内細胞密度および機能が厳密に制御されている4).特に,細胞寿命の長いミクログリアにおいては,ストレスに伴う細胞老化や細胞死,または細胞の若返りといった概念が重要な意味合いをもつ.本特集では,ミクログリアの細胞老化や若返りに関する最新の知見に関して紹介する(松平の稿).
2ミクログリアの機能や表現型を決定づける因子
個々のミクログリアの生存・成熟,もしくは表現型の決定付けには多くの因子が関与している.他のすべてのマクロファージと同様に,コロニー刺激因子1受容体(CSF1RもしくはCD115)を介したシグナルは,ミクログリアの発達,維持,活性化に必須である5).そのため,CSF1Rのシグナルを阻害すると,ミクログリアが脳実質内から消失する6).また,CSF1Rの遺伝的変異によって,多数の軸索腫大の出現を特徴とする白質脳炎が生じることが知られている7)8).脳内において,CSF1Rのリガンドとして知られるCSF1(M-CSF)およびインターロイキン34(IL-34)は,さまざまな細胞から供給されているが,それぞれのリガンドに対する依存度は,発達時期や脳領域によって異なる9).例えば,IL-34は灰白質領域のミクログリアの維持に重要な因子である一方で,白質領域のミクログリアはCSF1依存的である9)10).これら因子に加えて,トランスフォーミング増殖因子β(TGF-β)もミクログリアにとって重要な組織ニッチ形成分子である11)12).TGF-βは,神経細胞,アストロサイト,オリゴデンドロサイト前駆細胞,血管系細胞,そしてミクログリア自身が産生・放出すると考えられている13).実際,中枢神経系におけるTGF-βをノックアウトすることで,ミクログリアの減少と表現型の異常が引き起こされる11).特に,正常ミクログリアの形成・維持に重要な役割を果たすことが知られる転写因子Sall1の発現減少を伴って,形態学的な変化や正常細胞マーカー分子の発現低下,サイトカインの放出増加が引き起こされることを考慮すると14),TGF-βは正常脳内におけるミクログリアの機能維持に重要なシグナル分子であることは明白である.
こうした脳実質内から供給されるシグナル分子に加えて,遠位のミクログリア制御因子として腸内細菌叢が知られている15).腸内細菌から放出される短鎖脂肪酸は,ミクログリアの成熟に必須であり,これら代謝産物のシグナルを受けないミクログリアは,細胞内代謝に変化が生じ,またより反応性の高い未成熟な表現型を呈するようになる16).さらに,老齢マウス脳内で観察されるミクログリアの活性化は,若年マウスの腸内細菌を移植することで抑制される.腸内細菌がいかにして(直接的か,間接的か)ミクログリアの機能・表現型を制御しているのかについては議論の余地が残っているが,前記の報告は,腸内細菌叢の変化,つまりは腸内環境がミクログリアの状態変化と密接にかかわっていることを意味している.
3ミクログリアの機能と多様性,および中枢神経系疾患への関与
ミクログリアは,脳内に定着した直後からさまざまな機能を発揮する.発達期の脳内では,過剰な神経の除去や血管新生,オリゴデンドロサイト前駆細胞の保護や神経新生の促進,シナプスの剪定など多岐にわたる17).また成体脳内においては,常時細胞突起を使い,周囲環境を監視している.正常時には,常時伸縮をくり返しているが,突起伸展の方向性は必ずしもランダムに起きているわけではなく,特定の神経の細胞体やシナプスを標的としている可能性もある.特に,突起の接触頻度や接触時間に依存して神経の機能変容や神経シナプスの形成もしくは剪定が促進されることを考慮すると,ミクログリアが神経活動を積極的に制御していると考えられる.その他,神経保護やミエリンの機能維持など,さまざまな機能を担うことが明らかになっているが,本特集ではミクログリアと血液脳関門の透過性制御について紹介する(和氣の稿).
正常時に加え,ミクログリアはさまざまな中枢神経系疾患の発症に関与することが明らかになっている.アルツハイマー型認知症やパーキンソン病,筋委縮性側索硬化症や多発性硬化症といった神経変性疾患,てんかん発作,脳梗塞,脳腫瘍,精神疾患や神経障害性疼痛など,今では中枢神経系がかかわるほぼすべての疾患に関与するとまで言われている18).しかし,疾患時にミクログリアが担う役割,つまり病態発症メカニズムは多様で,サイトカイン等の液性因子の放出による炎症状態の惹起や,神経系の機能変容,神経シナプスの貪食の増減など,ミクログリアは異なるメカニズムで各疾患にかかわっている.本特集では,ミクログリアが関与する中枢神経系疾患の例として,ミクログリアによる神経回路編成とてんかん発作(安藤・小山の稿),および神経障害性疼痛発症・回復におけるミクログリアの役割(津田の稿)に関して概説する.
ところで,ミクログリアは,いかにして多種多様な機能を全うしているのか? その答えの一つでもあり,最近のホットトピックとなっているのが「多様性」である.神経系の研究領域では決して真新しいトピックではないが,ミクログリアの多様性に関する議論には長い歴史があり,細胞形態や分布密度,免疫関連分子等の細胞膜上発現分子パターンといった指標を用いた細胞の機能的分類では包括的な細胞分類が不可能ではないかと考えられてきた19).こうした議論に革新をもたらしたのが,最近急速な進歩を遂げているシングルセルRNAシークエンシング(scRNA-seq)を含めた1細胞解析技術である.1細胞レベルで得られた多次元情報の統合・比較解析は,細胞の多様性に関する議論にブレークスルーをもたらしている.本トピックに関しての詳細は,本誌・実験医学増刊「神経免疫 メカニズムと疾患〜神経系と免疫系を結ぶ分子機構の解明からバイオマーカー・治療標的の探索まで」(Vol.39-No.15,2021)をご覧いただきたい.
4ミクログリアの機能解析を目的とした研究ツール
これまで,エレクトロポレーション法などの古典的な遺伝子導入手法に加え,アデノ随伴ウイルス(AAV)ベクターやレンチウイルスを用いたミクログリアへの遺伝子導入が試みられてきた.それらは,培養細胞レベルでは一定の導入効率を達成し,ミクログリアの細胞機能の一端を明らかにしてきた.しかし,ウイルスベクターを用いた実験では,ベクターそのものによる免疫応答が誘導されることも多く,またin vivoでの導入効率の低さが問題となっている.そういった経緯もあり,これまでミクログリア研究を支えてきたのが遺伝子改変マウス等の実験動物である.実際,フラクタルカイン受容体(Cx3cr1)やCsf1rを標的とした遺伝子改変マウスは,ミクログリア研究のゴールドスタンダートとして,多くの革新的な発見に寄与してきた20).最近では,より細胞種特異性を考慮した遺伝子改変マウス(P2ry12CreERT2,Tmem119CreERT2,HexbCreERT2)も開発されており21)〜23),ミクログリア研究がこれまで以上に深化していくものと期待される.つい最近,AAVベクターを用いた遺伝子導入においても,その導入効率や特異性が格段によくなった方法が報告され24),より簡便かつ短時間でin vivoでのミクログリアの機能や特性を明らかにできるようになるかもしれない.
5ミクログリアと他の脳内免疫細胞
脳内の主要免疫細胞として認識されてきたミクログリアに加えて,髄膜(硬膜・くも膜・軟膜)や血管周囲スペース,脈絡叢といった脳境界領域には,脳境界マクロファージとよばれる細胞集団が存在しており,それぞれ重要な役割を果たしている25).特筆すべきは,ミクログリアと脳境界マクロファージが非常に似通った細胞特性を有している点である.特に,ミクログリアのマーカー分子として頻用されているIba1やCx3cr1は,同時に脳境界マクロファージにも発現しているため,これまで両者を正確に分けて解析した報告がきわめて少ない.しかし,ミクログリアの機能を正確に理解するためには,無視することのできない存在として認識されるようになり,ようやく脳境界マクロファージに関する理解が進み始めた.本特集では,脳境界マクロファージに関してミクログリアとの類似点や相違について最新の知見を交えて概説する(各論の増田の稿).
一方,免疫特権を有した組織とされてきた中枢神経系において,存在しえないと考えられてきた獲得免疫系細胞も多く存在することが明らかになっている.特にT細胞やB細胞などは,正常時から主に硬膜周辺に存在しており,脳内免疫に積極的に関与している26).さらに,頭蓋骨の骨髄内と脳硬膜を直接接続するチャネルの存在が明らかになり,じつは頭蓋骨から効率的かつ即時的に脳内に免疫細胞が動員できるシステムが存在していたのである27).こうした最近の動向に加え,中枢神経系疾患時における獲得免疫系細胞の関与について紹介する(伊藤の稿).
おわりに
近年,研究技術の革新により,ミクログリアの細胞特性に関する理解が急速に深化し,またミクログリアはさまざまな中枢神経系疾患に対する有望な治療標的として認識されてきた.ヒトミクログリアに関する情報が乏しいことなど,いまだ解決すべき点は多いが,今後動物モデルからヒトへの橋渡し研究が加速され,ミクログリアを標的とした疾患治療法が確立されることを期待したい.
文献
- Geirsdottir L, et al:Cell, 179:1609-1622.e16, doi:10.1016/j.cell.2019.11.010(2019)
- Prinz M, et al:Cell, 179:292-311, doi:10.1016/j.cell.2019.08.053(2019)
- Kierdorf K, et al:Nat Neurosci, 16:273-280, doi:10.1038/nn.3318(2013)
- Askew K, et al:Cell Rep, 18:391-405, doi:10.1016/j.celrep.2016.12.041(2017)
- Stanley ER & Chitu V:Cold Spring Harb Perspect Biol, 6:a021857, doi:10.1101/cshperspect.a021857(2014)
- Green KN, et al:Trends Immunol, 41:771-784, doi:10.1016/j.it.2020.07.001(2020)
- Nicholson AM, et al:Neurology, 80:1033-1040, doi:10.1212/WNL.0b013e31828726a7(2013)
- Rademakers R, et al:Nat Genet, 44:200-205, doi:10.1038/ng.1027(2011)
- Easley-Neal C, et al:Front Immunol, 10:2199, doi:10.3389/fimmu.2019.02199(2019)
- Kana V, et al:J Exp Med, 216:2265-2281, doi:10.1084/jem.20182037(2019)
- Butovsky O, et al:Nat Neurosci, 17:131-143, doi:10.1038/nn.3599(2014)
- Zöller T, et al:Nat Commun, 9:4011, doi:10.1038/s41467-018-06224-y(2018)
- Vanlandewijck M, et al:Nature, 554:475-480, doi:10.1038/nature25739(2018)
- Buttgereit A, et al:Nat Immunol, 17:1397-1406, doi:10.1038/ni.3585(2016)
- Erny D & Prinz M:Glia, 68:1655-1672, doi:10.1002/glia.23822(2020)
- Erny D, et al:Cell Metab, 33:2260-2276.e7, doi:10.1016/j.cmet.2021.10.010(2021)
- Prinz M, et al:Annu Rev Immunol, 39:251-277, doi:10.1146/annurev-immunol-093019-110159(2021)
- Priller J & Prinz M:Science, 365:32-33, doi:10.1126/science.aau9100(2019)
- Ydens E, et al:Nat Neurosci, 23:676-689, doi:10.1038/s41593-020-0618-6(2020)
- Yona S, et al:Immunity, 38:79-91, doi:10.1016/j.immuni.2012.12.001(2013)
- Masuda T, et al:Nat Immunol, 21:802-815, doi:10.1038/s41590-020-0707-4(2020)
- McKinsey GL, et al:Elife, 9:e54590, doi:10.7554/eLife.54590(2020)
- Kaiser T & Feng G:eNeuro, 6:ENEURO.0448-18.2019, doi:10.1523/ENEURO.0448-18.2019(2019)
- Lin R, et al:Nat Methods, 19:976-985, doi:10.1038/s41592-022-01547-7(2022)
- Kierdorf K, et al:Nat Rev Neurosci, 20:547-562, doi:10.1038/s41583-019-0201-x(2019)
- Alves de Lima K, et al:Annu Rev Immunol, 38:597-620, doi:10.1146/annurev-immunol-102319-103410(2020)
- Cugurra A, et al:Science, 373:eabf7844, doi:10.1126/science.abf7844(2021)
本記事のDOI:10.18958/7149-00001-0000279-00
著者プロフィール
増田隆博:2011年九州大学大学院にて学位取得・博士(薬学).九州大学大学院薬学研究院 学術研究員,同 特任助教を経て,同薬学研究院ライフイノベーション分野助教.’15年からドイツ・フライブルク大学病院に留学(日本学術振興会海外特別研究員).’17年から同大学病院博士研究員.’20年九州大学大学院薬学研究院薬理学分野助教を経て,’21年より現職(准教授).










