
概論
最近のMYC研究の世界の動向をふまえて
Recent remarkable progress in the molecular bases of multifunctional MYC protein
奥田晶彦
Akihiko Okuda:Division of Developmental Biology, Research Center for Genomic Medicine, Saitama Medical University(埼玉医科大学ゲノム医学研究センター発生・分化・再生部門)
c-Mycは細胞の活発な細胞増殖因子や,嫌気的代謝促進因子としてがん細胞の特質に大いにかかわっている遺伝子であり,その遺伝子産物であるc-MYCタンパク質はパートナー因子であるMAXと相互作用することで転写因子として機能する.本稿では,MYCの特集の巻頭として,c-Mycのバーキットリンパ腫の原因遺伝子としての発見から,c-Mycがきわめて多種多様な組織の腫瘍化にかかわることを可能にする分子基盤の解明,最近のトピックに至るまでのMYC研究の歴史について概説する.
はじめに
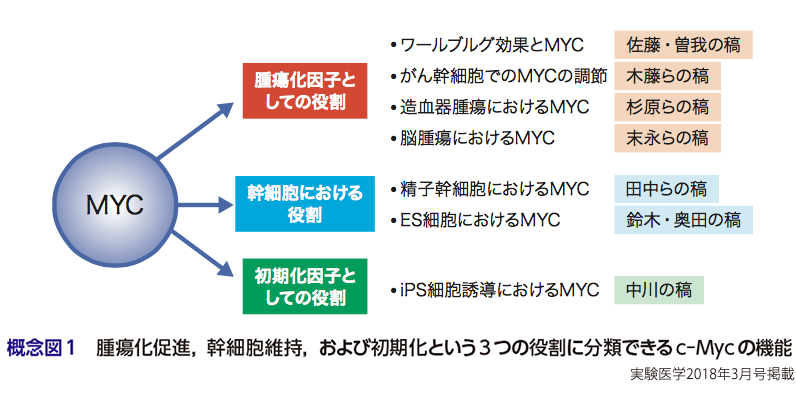
MYC,別名c-Mycのcはcellularのcであり,46年前にバーキットリンパ腫の原因遺伝子として,トリ白血病ウイルスがもつv-Myc遺伝子の細胞性のカウンターパートして同定された遺伝子である1).そして,その報告では,c-Myc遺伝子が染色体転座により,イムノグロブリン重鎖遺伝子のエンハンサーにより制御されるようになったことでB細胞において過剰発現し,そのことがこのリンパ腫の原因になっていることが示されている.それ以降,多くの種類のがんにおいて,MYC(c-Mycと同じファミリーに属するN-MycやL-Mycも含む)タンパク質の安定性が向上する変異であるとか,遺伝子増幅による発現上昇などが認められている2).このようにc-MYCは,古くから,腫瘍化に深くかかわる遺伝子として知られており,また,10年くらい前から,幹細胞維持にもかかわっていることや,iPS細胞誘導のための初期化因子であることが示され,現在,そういった方面の研究もさかんに行われている(概念図1).
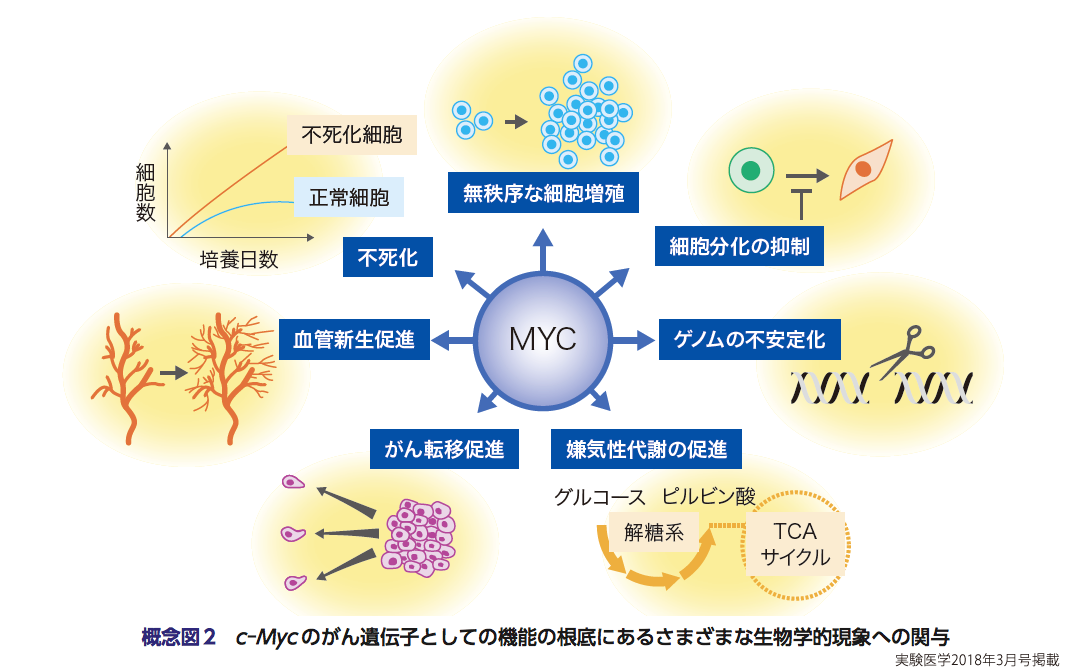
さらには,細胞増殖促進,代謝経路の変更など,c-Mycによる腫瘍化と,幹細胞維持,初期化といったMycの大きな3つの機能の根底にある分子基盤についての研究(概念図2)が近年,目覚ましい発展を示しているので,本稿では,MYCの特集の概論として,MYCの過去から最近のトピックに至るまでの研究について紹介することにする.なお,「MYC」と表記すると一般的にc-Mycを意味することが多いが,本稿ではMYCファミリーの総称として「MYC」を用いる.
1MYCの転写因子としての機能
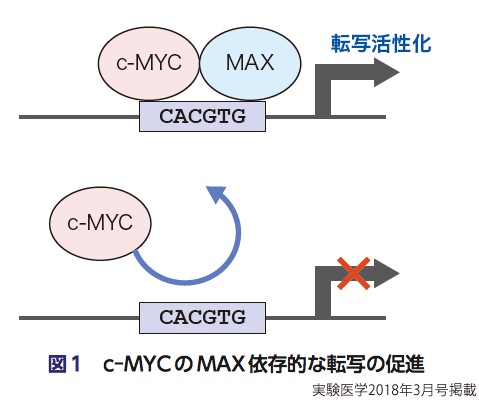
v-Myc遺伝子の発見に引き続き,c-Mycがバーキットリンパ腫の原因遺伝子として発見された事実は,MYC研究の歴史に最初に訪れた最も大きなブレークスルーであり,それについて語らずしてc-Mycの歴史を語ることができないが,それらc-Mycのがん遺伝子としての生物学的機能については,他の稿(杉原らの稿,および末永らの稿)で比較的詳しく記載されているので,それらの稿を参照されたい.c-Myc研究の歴史のなかでその次に訪れたブレークスルーは,c-MYCタンパク質が,がん遺伝子産物として機能するうえで必須なパートナー因子であるMAXタンパク質のクローニングであろう3).実際,MAXのクローニングの前から,MYCが転写因子として細胞の腫瘍化にかかわっているであろうことは想定されていたが,c-MYC (N-MYCもL-MYCもそうだが)は,MAXとの相互作用がなければ,転写因子として機能する以前にEボックスという特定の配列(5′-CACGTG-3′)に結合することすらできない(図1).したがって,MAXがクローニングされたことで,c-MYCが転写因子であることが証明され,そのおかげで,c-MYCの腫瘍化促進という生物学的な機能の根底を支えている分子基盤の解明が一気に進んだ.
2MYC自体の安定性についてのさまざまな調節
MYC自体,転写レベルでの調節に加え,RNAおよびタンパク質の安定性での調節といったように,さまざまなレベルでの調節を受けている.まず,転写レベルでの調節では,Myc遺伝子プロモーター上にBRD4という転写調節因子が結合し,転写を促進していることがわかっており,それゆえ,小分子化合物を用いてBRD4の活性を抑制することでMYCの発現を低下させ,がんを治療しようとする試みがなされている(杉原らの稿)4).転写されたMYCのmRNAレベルでの調節も知られている.実際,MYC mRNA自体,かなり不安定であるので,Myc遺伝子が強く転写されたとしても,細胞内でMYC mRNAは蓄積しがたく,さらには,MYC mRNAは,let-7であるとか,いくつかのmiRNAによる翻訳の抑制も受けている.ただし,MYC分子の調節という点で最も大きな要素は,58番目のスレオニン残基のGSK-βによるリン酸化であり,MYCタンパク質がこのリン酸化を受けると,プロテオソーム分解系によりすみやかに分解されることになる5).このようにc-MYCタンパク質は,正常細胞では,細胞内にあまり多く蓄積しないようにさまざまな調節を受けており,がん細胞は,そういったc-MYCタンパク質の量を制限する機構から回避することで,細胞内に比較的多くの量のc-MYCタンパク質の蓄積を達成している.その最も典型的な例としては,がん細胞が,リン酸化されないように58番目のスレオニン残基を変異させることで,c-MYCタンパク質の安定性を増していることがあげられる.なお,この調節については,杉原らの稿,佐藤・曽我の稿,および木藤の稿を参照されたい.
3c-Mycの嫌気的代謝促進因子としての役割
がん細胞は,一般的に,ワールブルグ効果といって,十分に酸素が得られる状況でも,エネルギー源としてミトコンドリアによる酸化的リン酸化反応(TCAサイクル)によりATPを得るのではなくて,酸素を必要としない解糖系代謝経路により主にATPを得ている(佐藤・曽我の稿).解糖系によりATPを得る方法は,TCAサイクルよりはるかに効率が悪いので,がん細胞は,大量のグルコースを消費する必要性が生ずる.がん細胞が,もっぱら解糖系によりATPを得ることの明確なメリットの一つは,解糖系により,グルコースから乳酸へと代謝される過程の最初の中間代謝産物であるグルコース 6-リン酸が,ヌクレオチドの合成を司るペントースリン酸経路の出発材料になることである.すなわち,活発な細胞増殖(DNA複製)のためのヌクレオチドを合成する必要のあるがん細胞にとって,グルコース 6-リン酸を比較的容易に利用できる環境は好都合なわけである.なお,このワールブルグ効果をがん細胞に賦与するうえで重要な働きをしているのがc-Mycであり,c-Mycは,グルコースの取り込みを司るGlut1遺伝子の発現や,グルコースからグルコース 6-リン酸への変換を触媒するhexokinase 2をコードする遺伝子などの発現を高めることで,この効果を発揮している6).なお,TCAサイクルは,ATPを得るためだけのものではなく,その中間代謝産物は,脂肪酸の合成など,いくつかの分子の合成のための出発材料として使われているので,がん細胞は,TCAサイクルを完全に止めるわけにはいかず,その問題点を克服するために,c-Mycを用いてグルタミンの取り込みを促進し,グルタミン酸を介してTCAサイクルのなかの一つの中間代謝産物であるα-ケトグルタル酸をつくることで,そこからのTCAサイクル経路を動かしている7).
4ES細胞維持因子および初期化因子としてのMycの役割
c-Myc,N-Myc,L-Mycはいずれもがん遺伝子として同定された遺伝子で,今でもがん研究との関連で最も研究されている遺伝子であるが,21世紀になってから間もない頃,ES細胞の未分化性維持において重要な働きを示すことが示された8)(鈴木・奥田の稿).そして現在では,造血幹細胞や精子幹細胞など,さまざまな組織幹細胞でのMycの重要性が示されている(田中らの稿).また,このMycのES細胞における重要性が示された研究成果の報告がきっかけとなって,MYCが初期化因子としての候補の一つとして考えられるようになり,そのことが,iPS細胞という,とてつもない偉業へとつながった9)(中川の稿).なお,ES細胞は,活発な細胞増殖など,さまざまな点においてがん細胞との類似性が指摘されているが,MYCがこれら2種類の細胞間における類似性の根底を支えている最も重要な責任分子の一つであることが示されている10).
5c-MYCの転写因子としての特殊性①
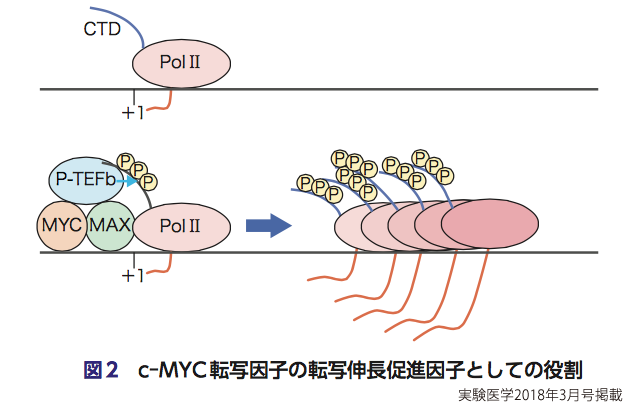
いわゆる一般的な転写因子の多くは,RNA ポリメラーゼⅡを遺伝子プロモーター上にリクルートし,転写を開始させることで転写を促進する.ただし,転写調節には,この転写開始促進という調節の他に,転写伸長というもう一つ重要な調節がある11).なぜならば,かなりの遺伝子において,RNAポリメラーゼⅡが転写のために,転写開始点から遺伝子上を滑り出したとしても,20~50塩基あたりのところでその動きを止めてしまうので,転写として遺伝子全体におよぶ機能的なRNAをつくるためには,いったん止まってしまったRNAポリメラーゼⅡを再び動かせることが必要になる.そして,c-MYCは,通常の転写因子とは違って,RNAポリメラーゼⅡを遺伝子プロモーター上によび込むのではなくて,もっぱら転写伸長を促進することで,転写を促進する転写因子であることが明らかにされた12).なお,このc-MYCは,RNAポリメラーゼⅡのC末端領域をリン酸化し,転写伸長を司るP-TEFbをリクルートすることにより転写伸長促進活性を発揮している(図2)
6c-MYCの転写因子としての特殊性②
c-Mycが高発現することで治療抵抗性を獲得するであるとか,がんとしての悪性度と深く関係していることはよく知られている事実であるが,これらc-Mycの発現量の変化に伴ってみられるがん細胞としての特質の変化は,c-Mycが低発現状態では発現がほとんどOFFの状態にある特定の遺伝子セットに,c-Mycが高発現することによって結合するようになり,そのことで,それらの遺伝子の発現がONになるのであろうと考えられていた.それゆえ,多くの研究グループが,そのようながん細胞の悪性化にかかわっているc-MYC標的遺伝子を同定すべく,さまざまながん細胞を用いてc-MYC転写因子のゲノムでの結合部位を網羅的に調べた.その結果,予想外なことに,それぞれのがん細胞において,c-MYCが結合しているゲノムに位置は,細胞増殖促進や代謝にかかわる遺伝子などのいくつかの遺伝子において共通していることを除き,ほとんど共通点がないことがわかった.そして,2つの米国のグループはこの事実に興味をもち,深く解析することで,c-MYCによる細胞の腫瘍化のためのたいへん重要な分子メカニズムを解明した13)14).これら2つのグループは,c-MYCがいわゆる特定の遺伝子セットの発現のON-OFFをコントロールするのではなくて,高発現したc-MYCは,c-MYC高発現前(c-MYCは低レベルで発現)においてもある程度のレベルの発現がみられる遺伝子に限って発現レベルを高め,がん細胞の特質を変えていることを証明した.この現象の根底にある分子メカニズムは,通常,c-MYC標的遺伝子は,c-MYCの発現レベルが低くても結合可能な完全なEボックス配列に加え,不完全なEボックス配列ももち,そこにはc-MYCが高発現しているときのみ結合が可能であるため,c-MYCの発現量の違いにより,標的遺伝子の発現レベルが変化するというものである.つまり,ON-OFFのスイッチではなく,ボリュームのコントローラーのようなはたらきだったのだ.
おわりに
c-Mycがバーキットリンパ腫の原因遺伝子であることが明らかになった40数年前から現在までのc-Myc研究の歴史について概説するつもりであったが,c-MycによるmicroRNA産生の抑制15)といったc-MYCの重要な機能であるとか,c-Mycの発現量を低く保つことで長寿を達成するというたいへん話題を集めた報告16)であるとか,c-Mycについての総説として当然紹介しなくてはならないことをいくつも,誌面の関係で記載することができなかった.その点については,少し心残りではあるが,この概説とc-Mycについての7つの各論を読んでいただければ,現在においてもMYC研究からきわめてインパクトのある新事実がどんどん解き明かされており,MYC研究が今でもきわめてホットな領域であることをわかっていただけるのではないかと期待している.
文献
- Taub R, et al:Proc Natl Acad Sci U S A, 79:7837-7841, 1982
- Dang CV:Cell, 149:22-35, 2012
- Blackwood EM & Eisenman RN:Science, 251:1211-1217, 1991
- Abedin SM, et al:Onco Targets Ther, 9:5943-5953, 2016
- Yada M, et al:EMBO J, 23:2116-2125, 2004
- Hu S, et al:Cell Metab, 14:131-142, 2011
- Wise DR, et al:Proc Natl Acad Sci U S A, 105:18782-18787, 2008
- Cartwright P, et al:Development, 132:885-896, 2005
- Takahashi K & Yamanaka S:Cell, 126:663-676, 2006
- Kim J, et al:Cell, 143:313-324, 2010
- Core LJ & Lis JT:Science, 319:1791-1792, 2008
- Rahl PB, et al:Cell, 141:432-445, 2010
- Lin CY, et al:Cell, 151:56-67, 2012
- Nie Z, et al:Cell, 151:68-79, 2012
- Chang TC, et al:Nat Genet, 40:43-50, 2008
- Hofmann JW, et al:Cell, 160:477-488, 2015
著者プロフィール
奥田晶彦:1984年,広島大学医学部医学科卒業,’88年,東京大学医学部博士課程修了,同年より日本学術振興会特別研究員,’90年から米国Caltechに留学,’92年,埼玉医科大学講師,2001年,埼玉医科大学ゲノム医学研究センター助教授(PI),’05年,同研究センター教授,’17年,同研究センター所長,現在に至る.埼玉医科大学に赴任以来,ES細胞研究を行っており,最近の10年間は,c-MYCを手掛かりに,ES細胞とがん細胞の類似性を規定している分子基盤の解明を主な目的として研究を行っている.










