概論
特集1 概論
獲得免疫を軸に免疫老化を解き明かす
Deciphering immunosenescence rooted in adaptive immunity
濵﨑洋子
Yoko Hamazaki:京都大学iPS細胞研究所未来生命科学開拓部門/京都大学大学院医学研究科免疫生物学
高齢者が特にハイリスクとなったCOVID-19パンデミックを機に,ヒト免疫の老化(加齢に伴う機能低下と制御異常)を理解する重要性が改めて認識され,近年大きな発展を遂げた高度オミクス解析技術により,その実態とメカニズム解明のための研究が急速に進んでいる.免疫老化は感染症だけでなく,がん・自己免疫疾患・代謝性疾患などさまざまな加齢関連疾患の発症と増悪の基礎となることから,免疫老化そのものを制御することによる加齢関連疾患の克服にも期待が集まっている.本特集では,主に獲得免疫システムを担う免疫細胞の老化を中心に,最近の知見を紹介する.
はじめに
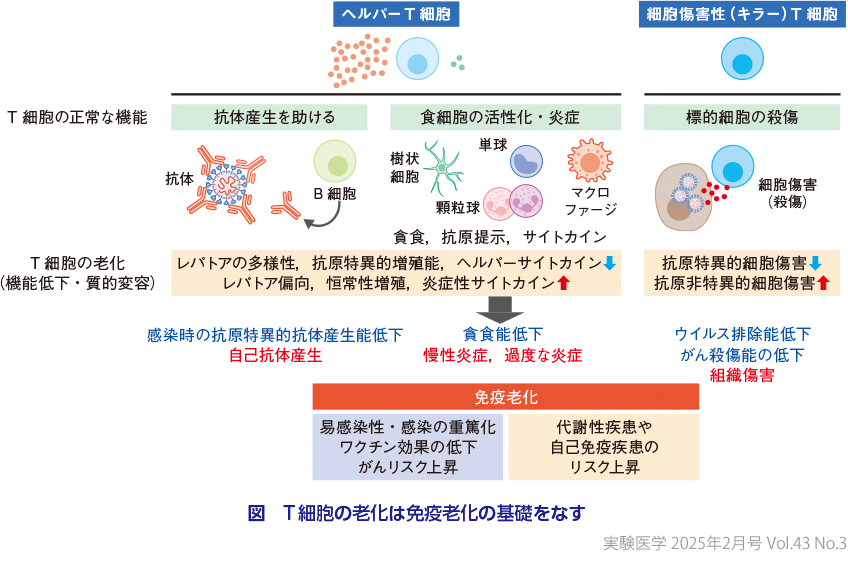
免疫老化は,加齢に伴い免疫システムの構成要素が数的・質的に変化し,われわれが期待する本来の免疫機能が低下する一方で,過度な炎症や感染のない状態での微弱な炎症が持続したり(慢性炎症),標的が本来攻撃すべきでない自己に向かったりするなど,免疫応答の制御異常(dysregulation)にある状態と定義される(図).免疫機能の低下は,易感染性や感染の重篤化,ワクチン効果やがんを排除する能力の低下につながりうる.加齢とともに細胞老化をきたしてSASP(senescence associated secretory phenotype,老化関連分泌表現型)を示す細胞が各臓器に蓄積する現象も,免疫系による老化細胞排除の効率低下が一因であるとされる.他方,過度な炎症は,COVID-19でも着目されたサイトカインストーム1)がそうであったように,免疫機能低下とは別のメカニズムで感染症の増悪をもたらす.また,慢性炎症は代謝性疾患の基礎となる.遺伝的素因の寄与が大きいとされる自己免疫疾患においても,加齢は大きなリスク因子の1つであり,元来有していた遺伝的素因が,免疫老化による免疫システムの制御異常の過程で顕在化すると考えられる2).
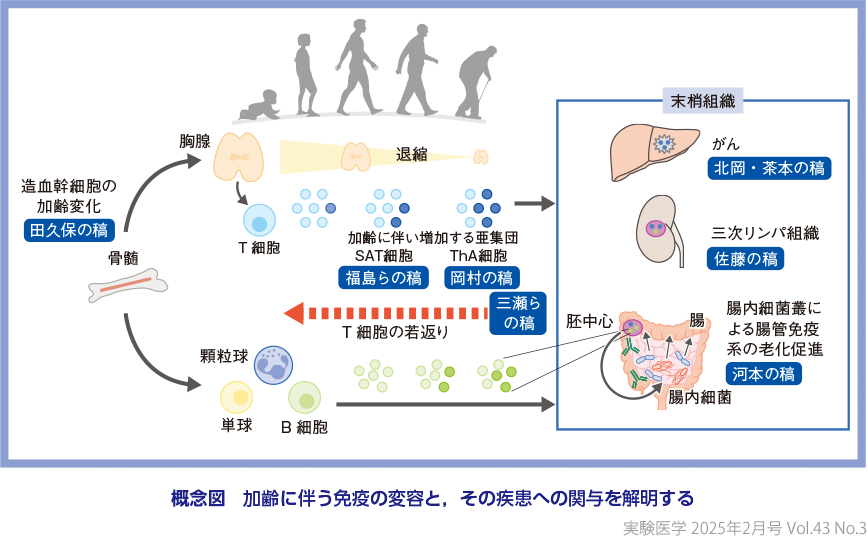
このように,免疫老化(機能低下と制御異常)はさまざまな加齢関連疾患の基礎をなすことから,各疾患に共通のリスク因子である免疫老化をターゲットに介入するという考え方も提案されている.しかしながら,単純な機能低下ではなく,好ましくない応答が増大する免疫老化の特性にかんがみると,その適切な制御が簡単でないことは容易に想像がつく.多種多様な機能をもつ免疫細胞が全身を駆けめぐり,非免疫細胞も含めたさまざまな組織の細胞と相互作用し,ブレーキとアクセルとの絶妙なバランスで成り立つ免疫システムならでは(しかし学問としては興味深い)といえるだろう.炎症老化(inflammaging)に関連するトピックスは最近の本誌特集にゆずり3),本特集では,主に獲得免疫細胞が主となる免疫老化の実態とそのメカニズムを中心に,それがどのように免疫システム全体の老化や加齢関連疾患にかかわるのか議論してみたい(概念図).
1ヒトとマウスにおける免疫老化の共通性
近年のオミクス解析技術の飛躍的進歩により,マウスのみならずヒト免疫細胞における加齢変化の実態が,単一細胞・分子レベルで詳細に記載できるようになった.得られるデータがきわめて拡大深化する一方で,in vivoにおけるメカニズムの検証や証明には課題もある.また,種を超えて保存されたメカニズムの解明や介入方法の探索には依然としてモデル動物の有用性も高い.このことから,まずはヒトとモデル動物として用いられるマウスで共通している特徴について,大局的に考えてみたい.
① 造血幹細胞の老化
造血臓器である骨髄において,造血幹細胞(hematopoietic stem cell:HSC)から多数のステップを経て赤血球や血小板,免疫細胞がつくられる.免疫細胞はT細胞やB細胞などのリンパ球系と顆粒球やマクロファージなどの骨髄(ミエロイド)系に分類される.リンパ球系免疫細胞の産生が加齢に伴い減少する現象はよく知られているが,この点はヒトとマウスで共通した現象である.本特集では,HSCの加齢変化について,代謝に着目したご自身の最新研究も含めて田久保圭誉先生に解説いただいた(田久保の稿).最近では,DNMT3A,TET2,ASXL1などのエピジェネティック修飾(制御)因子のHSCにおける変異がクローン性造血を引き起こすこと,さらにこれらの遺伝子の機能不全が,ミエロイド系に偏向した造血や,自然免疫系細胞の炎症性サイトカイン産生能亢進を介して,心血管疾患のリスクに関与することが明らかになった4).さらに,HSCの加齢変化に伴うミエロイド系細胞の形質変化が,がん微小環境を修飾するとの報告も最近なされている(北岡・茶本の稿).シングルセル解析を用いてマウスとヒトの加齢に伴う免疫細胞のサブセット変化を比較した研究においても,ある程度の類似性があることが報告されている5).例えば,形質細胞様樹状細胞の加齢に伴う減少も共通しているが,このことは,COVID-19でも着目されたウイルス感染の初期応答に重要なtypeⅠIFN産生能の低下が,加齢に伴う感染に対する脆弱性の上昇を説明する種を超えた一因であることを示唆している.
② 加齢の影響を受けやすいT細胞
T細胞は,心臓の上に位置する臓器である胸腺の中で分化・選択を受けたのち,全身に分布する.その機能や分化段階から,多数の亜集団(サブセット)に分けられる.さまざまな種類の免疫細胞のなかで特にT細胞において加齢に伴う数的・質的変化が顕著であるという点も種間で共通した特性である(図).原因の1つに,T細胞産生臓器である胸腺の早期退縮があげられる.その結果,多様な未知の抗原への反応性を担保するナイーブT細胞が加齢に伴って減少する.顕著に低下しつつも残存する成人期の胸腺の活動の機能的意義を明確に示すエビデンスは乏しかったが,最近,成人で胸腺摘出を伴う治療を受けた患者において,全死亡率や,がん・自己免疫疾患の発症リスクが有意に高く,血中サイトカインがよりproinflammatory(炎症促進的)なパターンを示すことが示された6).この現象が,ナイーブT細胞の供給低下に依存するのか,免疫抑制機能を有する制御性T細胞(Treg)の産生低下に関連するのか,現時点で定かではないが,この最新の報告によって,ヒトの長い一生でT細胞と免疫システムの恒常性維持に退縮後の胸腺がどのように寄与しているのか,近年注目が集まっている.
こうした知見は,成獣マウスから胸腺を摘出し新たなT細胞産生を遮断すると,恒常性増殖が促進され,加齢で増加する老化関連T細胞(senescence-associated T cell:SAT細胞)が早期に誘導されるという現象とも矛盾しない(福島らの稿).われわれも以前に,胸腺摘出によりT細胞の老化を促進させる本モデルを用いて,加齢で増加し抗原刺激に対して迅速にIFNγを産生するCXCR3+ ナイーブCD8+ T細胞亜集団を同定した7).重要なことに,CXCR3+ ナイーブCD8+ T細胞は,末梢血ヒト検体を用いた最新のシングルセル解析技術を用いて,ヒトにおいても加齢で増加するユニークな集団として最近同定されている8).さらに,supercentenarian(110歳以上の高齢者)で増加するサブセットとして注目される細胞傷害性CD4+ T細胞(CD4 CTL)や,細胞老化をきたした組織の細胞からのSASP産生を促進するとされるGZMK+ CD8+ エフェクターメモリーT(Tem)細胞も,加齢とともに両種において蓄積する9)10).また,過度な免疫応答を抑制するTregが,両者で加齢により増加し機能も亢進しているとの報告もある.組織炎症を抑えるために代償的に増え,そのために正常な免疫応答が低下するとの示唆もあり,抗原特異性については議論は十分になされていないものの,免疫老化を制御する重要なプレーヤーであると考えられる11)12).
これらのことから,寿命や生活・微生物環境の違いや,ヒトで古くから知られているT細胞表現型の加齢変化(CD27/CD28陰性分画の増加,CD56/CD57陽性分画の増加など)がマウスでは認められないといった違いはあるものの(岡村の稿),免疫老化をもたらすメカニズムのなかには,種を超えた共通のメカニズムが存在することが示唆される.
2ヒトT細胞老化の特徴とその要因
① CD8+ T細胞の加齢変化が特に顕著である
末梢血中のCD8+ T細胞における加齢に伴うナイーブ型の割合および数の減少は,ヒトで特に顕著である13)14).COVID-19においては,ナイーブCD8+ T細胞の割合が低いことと重症化の関連が強く示唆されており15),多様な反応性を有し未経験の抗原に対する反応性を担保するナイーブT細胞の機能的重要性を示唆しているものと考えられる.われわれが最近行った,COVID-19 mRNAワクチン後の抗体応答の予測因子を同定する多変量解析を用いた探索的研究においても,ナイーブCD8+ T細胞の割合が,最も重要な独立予測因子として同定された16).本来CD4+ T細胞が関与する抗体応答を目的因子としたため,CD8+ T細胞のパラメーターが予測因子として同定されたことは意外であったが,加齢変化の大きいナイーブCD8+ T細胞の割合が,免疫機能全体の年齢(一般に言う “免疫力” や “免疫年齢” )の指標の1つとなりうる可能性を示唆しており興味深い.
ナイーブ分画の減少に伴い,高度に機能分化し増殖能を失った細胞や持続的な抗原刺激による疲弊や細胞老化(岡村の稿,北岡・茶本の稿,三瀬らの稿)をきたしたT細胞分画割合の増加も顕著であり,機能再生の重要な標的であるといえる14).茶本健司先生らには,質的変化の重要な側面であるミトコンドリア代謝調節異常を含めて,がん免疫における免疫老化の意義について解説いただいた(北岡・茶本の稿).また,吉村昭彦先生らには,T細胞が疲弊するメカニズムの研究から見出されたNR4aを標的としたT細胞若返りの試みについてご紹介いただいた(三瀬らの稿).
② CD4+ T細胞は数よりも質的変化が焦点
ヒトでは末梢での恒常性増殖(北岡・茶本の稿)により,CD4+ T細胞におけるナイーブT細胞の割合やT細胞抗原受容体(TCR)の多様性は比較的良好に長期維持される14).それでも,われわれが行ったCOVID-19 mRNAワクチン接種後の免疫応答解析においては,高齢者では抗原特異的T細胞の誘導が遅れかつ応答の収束が早期に起こることが示された17).さらに最近の研究で,ブースター接種後の記憶応答において,高齢者でもB細胞応答が増強され,成人群と同等の抗体が産生されるものの,T細胞応答は依然として低い傾向にあることが明らかになった18).このT細胞応答の質的変化のメカニズムは明らかではないが,高齢者ではTCRシグナル伝達が減衰しているにもかかわらず,抗原刺激によってエピゲノムのリモデリングとエフェクター細胞の分化を促進する転写因子ネットワークが誘導されやすいとの報告が最近なされており19),こうしたメカニズムと関連する可能性がある.
本特集では加齢に伴って増加するCD4+ T細胞亜集団として,SAT細胞,ThA細胞(age-associated helper T cell,加齢関連ヘルパーT細胞)について,それぞれ福島祐二先生ら,岡村僚久先生にご自身の研究を含めてご解説いただいている(福島らの稿,岡村の稿).これらの亜集団が,マウス(SAT細胞),ヒト(ThA細胞)で共通して加齢だけでなく自己免疫疾患や代謝性疾患でも増加することは,これらの疾患が確かに免疫老化と関連することを示唆しており興味深い.CD4+ T細胞が,抗体産生,貪食活性,炎症,細胞傷害活性など多様な免疫エフェクター機能を制御することを考えると,その機能的変容が免疫システム全体を制御異常に導くメカニズムが示唆される.さらに,岡村先生らが同定したThA細胞において,その機能的特性を特徴づける転写因子(ZEB2)が同定されたことは特筆すべきである(岡村の稿).一方で,健常人と自己免疫疾患患者のThA細胞は質的に異なることも示されており,各組織の状態や炎症に応答して機能を獲得しているものと推察される.加齢で増加する集団が次々と同定され,さらに大きな個人差を認めるなかで,それらをどのように定義・整理していくのかは今後の課題である.
3ヒト免疫老化の理解と制御に向けて
① ヒトの各臓器における免疫老化の実態と病態形成における意義
オミクス解析技術が進歩した今も,ヒトでは末梢血検体を用いた解析が主であり,実際の病態を呈する末梢臓器において,どのように免疫老化が病態を修飾しているのかを理解することは未解決の重要課題である.この点において,高齢者腎臓とヒト自己免疫疾患の末梢組織にみられる三次リンパ組織(tertiary lymphoid structure:TLS)に着目したユニークな研究を,佐藤有紀先生に紹介いただいた.マウスモデルとヒト検体,おのおのの強みを活用した研究としても示唆に富む内容である(佐藤の稿).マウスで実施されているさまざまな組織の免疫細胞シングルセルプロファイリング20)や近年急速な進化を遂げている空間トランスクリプトミクスの加齢変化解析も含め,免疫老化がどのようにヒト末梢組織での病態形成を時空間的に制御しているのか,実態解明が進むことが期待される.
② 免疫老化の個人差形成の要因とメカニズムは何か? 免疫年齢は定義できるか?
特定の病原体のない環境で生活している実験用マウスとは異なり,ヒトは感染によって常に免疫系が再構築されている.いくつかの潜伏ウイルス感染〔サイトメガロウイルス(CMV)感染など〕がヒト免疫老化と関連することはよく知られている.一方で,CMV感染は若齢者ではむしろ免疫賦活化に働きうること,無菌マウスは免疫システムが未発達であることを考えると,単純に感染の刺激が免疫老化を促進すると結論づけるのは拙速である21).CMV特異的T細胞のクローンサイズが高齢になると増大するなど,CMV感染がT細胞に大きな影響を及ぼすことは間違いないものの,最近の研究では,細胞老化によって皮膚線維芽細胞に潜伏感染したCMVが再活性化され,CMV由来の抗原を標的として前述したCD4 CTLが老化細胞を排除しうることが示された.こうした結果から,加齢に伴う老化細胞の蓄積を防ぐためにCMVとの共生関係を確立するよう,免疫系が進化したのではないかという興味深い仮説が提唱されている22).
同様の観点で,腸内細菌や皮膚・粘膜部位の常在菌は,個人差が大きく常に宿主を刺激しつづけ免疫システムを修飾する重要なプレーヤーである.細菌叢のバランスの乱れ(dysbiosis)が,炎症性疾患や老化を促進するとの概念が確立しており,2023年には「hallmarks of aging」にも追加された23).河本新平先生には,腸内細菌叢の長期的な刺激による胚中心B細胞の細胞老化がdysbiosisを起こすという新たなメカニズムについて解説いただいた(河本の稿).B細胞の老化という観点では,加齢で増加するage-associated B cell(ABC,加齢関連B細胞)が最近のトピックスであり,T細胞との関係も含めて岡村の稿,佐藤の稿で紹介いただいている(概念図).
最後に,老化を促進する遺伝的素因の存在が示唆されていることから24),免疫老化の個人差にも遺伝的素因が存在する可能性について触れておきたい.われわれのCOVID-19 mRNAワクチン接種後の免疫応答の解析からも,年齢差だけでなくHLA型の多型以外にも遺伝的素因が存在する可能性を示唆する大きな個人差が認められた17).自己免疫疾患解析で威力を発揮しているeQTL解析などによって,免疫老化を制御する免疫細胞と遺伝的素因が明らかになるものと期待される.
おわりに
オミクス解析技術の発展により,今後もヒト免疫細胞の加齢変化の実態が単一細胞レベルで解析され,より詳細に分子レベルで記載されるとともに,加齢で増加する機能的変容をきたした免疫細胞亜集団が同定されていくと考えられる.この膨大なデータを活用し,ヒト検体と動物モデルの共通点と相違点,強みと限界を理解し,おのおのを活かした補完的なアプローチを用いて,ヒト免疫老化を個体レベルで理解する方向性が今後ますます必要になると考えられる.さらに,免疫老化の個人差形成にかかわる要因とメカニズムを解明することは,多様な遺伝的バックグラウンドと環境要因の影響のなかで起こるヒトの加齢関連疾患の発症と病態を理解するうえでの大きな課題である.これらの知見をもとに,ヒト免疫老化の本質的理解と,加齢関連疾患の包括的予防・介入につながる方法論開発の実現が期待される.
文献
- 「実験医学 Vol.39 No.4 免疫系の暴走 サイトカインストーム」(村上正晃/企画),羊土社(2021)
- Ray D & Yung R:Clin Immunol, 196:59-63, doi:10.1016/j.clim.2018.04.002(2018)
- 「実験医学 Vol.41 No.19 炎症老化 Inflammaging」(尾池雄一,真鍋一郎/企画),羊土社(2023)
- Evans MA & Walsh K:Physiol Rev, 103:649-716, doi:10.1152/physrev.00004.2022(2023)
- Mogilenko DA, et al:Nat Rev Immunol, 22:484-498, doi:10.1038/s41577-021-00646-4(2022)
- Kooshesh KA, et al:N Engl J Med, 389:406-417, doi:10.1056/NEJMoa2302892(2023)
- Kato A, et al:Eur J Immunol, 48:1663-1678, doi:10.1002/eji.201747431(2018)
- Bohacova P, et al:Immunity, 57:2362-2379.e10, doi:10.1016/j.immuni.2024.08.019(2024)
- Hashimoto K, et al:Proc Natl Acad Sci U S A, 116:24242-24251, doi:10.1073/pnas.1907883116(2019)
- Mogilenko DA, et al:Immunity, 54:99-115.e12, doi:10.1016/j.immuni.2020.11.005(2021)
- Jagger A, et al:Gerontology, 60:130-137, doi:10.1159/000355303(2014)
- Palatella M, et al:Front Immunol, 13:940705, doi:10.3389/fimmu.2022.940705(2022)
- Jo N, et al:Front Aging, 2:719342, doi:10.3389/fragi.2021.719342(2021)
- Goronzy JJ & Weyand CM:Nat Rev Immunol, 19:573-583, doi:10.1038/s41577-019-0180-1(2019)
- Rydyznski Moderbacher C, et al:Cell, 183:996-1012.e19, doi:10.1016/j.cell.2020.09.038(2020)
- Hidaka Y, et al:in revision
- Jo N, et al:Nat Aging, 3:751, doi:10.1038/s43587-023-00426-w(2023)
- Kometani K, et al:Front Immunol, 15:1455334, doi:10.3389/fimmu.2024.1455334(2024)
- Zhang H, et al:Nat Immunol, 24:96-109, doi:10.1038/s41590-022-01369-x(2023)
- Krishnarajah S, et al:Nat Aging, 2:74-89, doi:10.1038/s43587-021-00148-x(2022)
- Jergović M, et al:Med Microbiol Immunol, 208:263-269, doi:10.1007/s00430-019-00605-w(2019)
- Hasegawa T, et al:Cell, 186:1417-1431.e20, doi:10.1016/j.cell.2023.02.033(2023)
- López-Otín C, et al:Cell, 186:243-278, doi:10.1016/j.cell.2022.11.001(2023)
- Melzer D, et al:Nat Rev Genet, 21:88-101, doi:10.1038/s41576-019-0183-6(2020)
本記事のDOI:10.18958/7657-00001-0001822-00
著者プロフィール
濵﨑洋子:京都大学大学院医学研究科月田承一郎教授のもと上皮細胞生物学を学び学位取得.胸腺上皮細胞に興味をもつようになり湊長博教授の研究室で胸腺発生研究を開始する.iPS細胞研究所で独立後は,ヒト免疫老化の研究に加えて,胸腺上皮細胞の成り立ちをより深く知り,ナイーブT細胞の再生による免疫老化の克服を夢見て,iPS細胞から胸腺上皮細胞を分化誘導する研究を行っている.シャーレの中で徐々に胸腺上皮細胞へと分化していく様を目の当たりにし,そのポテンシャルに今さらながらに感動している.
コロナ禍の挑戦:ワクチン応答解析でヒト免疫老化の実態に迫る
マウスモデルを用いてT 細胞産生臓器である胸腺の発生と退縮,T 細胞の老化研究を行いながら,ヒトでもみてみたいと若齢者と高齢者の末梢血検体を用いてT 細胞解析系を立ち上げつつあるなかで起きたCOVID-19 パンデミック.ワクチン接種がヒト免疫老化研究の絶好の機会だと気づき,京大病院の先生方に何とか採血に協力していただきたいとお願いして回り,健常者コホート研究が実現した.厳しい感染対策のなか,病院の医師・スタッフの方々,ラボメンバーと協力して研究を進められたことはすばらしい経験となった.ワクチンという同じ抗原刺激を受けた後の免疫応答に,年齢差のみならずあまりに大きな個人差が存在することに衝撃を受けるとともに,これがヒト免疫老化の実態だと実感した.採血に協力してくださっているボランティアの皆さんにも,この場をお借りして心から感謝したい.(濵﨑洋子)









