
概論
特集1 概論
拡大する鉄代謝とフェロトーシス研究
学際的なハブとしての新展開
Emerging roles of iron metabolism and ferroptosis: New frontiers as an interdisciplinary hub
諸石寿朗
Toshiro Moroishi:東京科学大学総合研究院難治疾患研究所細胞動態学分野
鉄は生命活動に必須の微量金属元素であり,細胞内で多くの酵素反応を調節する.近年,鉄毒性が引き起こす新たな細胞死のメカニズムとしてフェロトーシス(ferroptosis)が発見され,鉄代謝,脂質代謝,レドックス制御のバランスが崩れることにより細胞傷害が引き起こされることが明らかになった.このメカニズムは,がんや老化,組織傷害などの理解に新たな視点をもたらし,その予防法や治療法の開発への応用が期待されている.本特集では,フェロトーシスの分子機構やその生理的・病理的意義について最新の研究成果を紹介し,拡大するフェロトーシス研究の理解を深める.
はじめに
われわれヒトの体に,どれだけの量の鉄が含まれているかをご存知だろうか? その量は鉄釘1本分(約5 g)にも及ぶ.鉄はさまざまな生命活動に欠かせない機能を果たす微量金属元素であり,例えば赤血球において鉄がヘモグロビンの活性中心として酸素運搬に必須の役割を担っていることは周知の事実である.一方,鉄が果たすその他の多様な生命機能について詳しく説明できる人は少ないのではないだろうか.近年,鉄が関与する新たな細胞死の概念としてフェロトーシスが知られるようになり,生命科学研究における鉄の重要性が再認識されている.
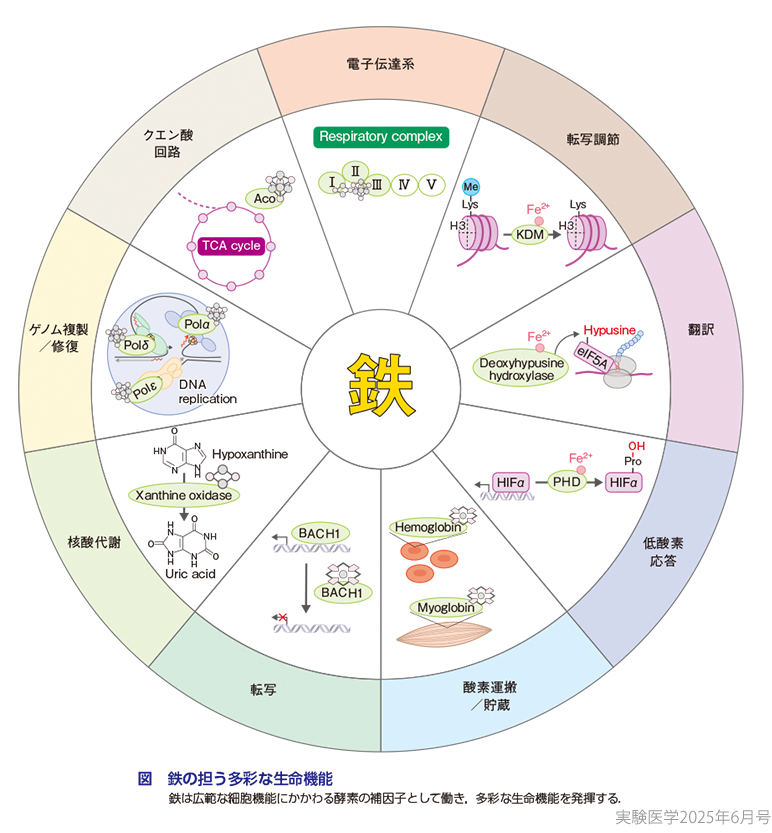
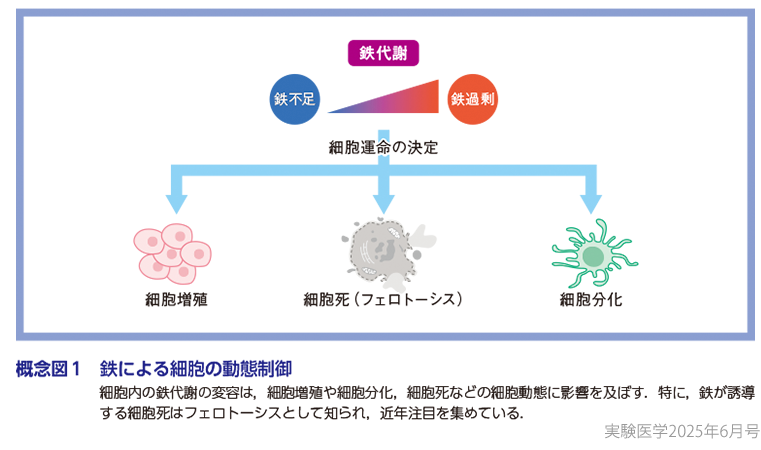
鉄は地球上に重量比で最も多く存在する元素であり,生命はその起源から鉄を利用して代謝活動を行い,生命を維持してきた.約40億年前に地球に誕生した,現存するあらゆる生命体の共通祖先であるLUCA(the last universal common ancestor)は,まだ地球上に酸素が存在しない時代に鉄を利用して嫌気的代謝を行い,その生命を維持していたと考えられている1).このように,鉄は電子を容易に授受しやすい化学的性質から多くの酵素反応の足場となり,ほとんどの生物,および,ほとんどの細胞にとって広範な生命機能に必須の役割を担う.われわれヒトにおいては,約400種類の鉄結合タンパク質が存在するとされており2),鉄はゲノム複製やエピジェネティクスを介した転写調節,電子伝達系など広範な細胞機能にかかわる酵素の補因子として生命活動に欠かせない元素である(図)3).また,近年これらの鉄結合タンパク質の鉄に対する親和性は,ものによって大きく異なることも明らかにされてきた.例えば,同じヒストン脱メチル化酵素であるKDM3AとKDM3Bは,ともに鉄をその酵素活性に必要とするが,両者の鉄への親和性は約40倍異なり(Km[Fe]の値はKDM3Aが〜2.6 μM,KDM3Bが〜100 μM),鉄への親和性の低いKDM3Bが介在するH3K9me2の脱メチル化反応は,細胞が鉄欠乏に陥るとその活性が大きく低下する4).すなわち,鉄は酵素反応に必須であるのみならず,その活性を動的に制御しうる因子としても再注目されている.このような背景から,細胞における鉄代謝の変容は,細胞の増殖・分化・死などの細胞運命決定に重要な役割を担うと考えられ,鉄による細胞の動態制御が近年さかんに研究されている(概念図1).
このように鉄は生命活動に必須の役割を担う一方で,過剰な細胞内の遊離鉄はフェントン反応[Fe2++H2O2→Fe3++OH−+OH・]を介して酸化傷害を引き起こすなど,細胞毒性を有する.例えば,全身に鉄が蓄積するヘモクロマトーシスでは,鉄の蓄積による組織傷害から,肝障害,糖尿病,心筋症などの臓器障害をきたす5).これまで鉄による細胞傷害の分子基盤は明確ではなかったが,2012年に細胞内の鉄過剰による脂質の過酸化が惹起する新しい細胞死の概念としてフェロトーシスが発見され6),鉄毒性の分子機構が次第に解明されつつある.フェロトーシスはがんや神経変性疾患,虚血再灌流傷害などの病態に深く関与することが示され,フェロトーシス研究はますます広がりを見せている.そこで,本特集ではフェロトーシス研究の第一線で活躍する研究者の方々に,フェロトーシスの分子基盤や生物学的意義,病態とのかかわりについて紹介いただく.
拡大するフェロトーシス研究
フェロトーシスの発見以来,細胞のフェロトーシス感受性について多くの研究が蓄積されてきたが,これまでに明らかにされたフェロトーシスの三大要素は,①鉄代謝,②脂質代謝,③レドックス制御に集約される(概念図2)7).鉄はトランスフェリンと結合して3価(Fe3+)の状態で細胞内に運ばれ,2価(Fe2+)に還元されて遊離鉄という反応性に富んだ鉄のプールを形成する.遊離鉄はミトコンドリアへと運ばれ,鉄硫黄クラスターやヘムなどの鉄補欠分子族へと代謝され,前述のようにさまざまな酵素の補因子として働くことにより細胞活動に必須の役割を担う.一方,過剰な遊離鉄は鉄毒性を示すため,細胞内の鉄代謝は厳密に調節されている8).細胞内鉄動態の破綻により遊離鉄が増加すると,フェントン反応を介して脂質過酸化が引き起こされ,フェロトーシスが誘発される(𥱋取・岩井の稿).この脂質過酸化は,細胞膜リン脂質中の脂肪酸組成に大きく影響される.すなわち,多価不飽和脂肪酸(polyunsaturated fatty acid:PUFA)はラジカルにより容易に酸化され連鎖的脂質過酸化反応の起点となる一方,一価不飽和脂肪酸(monounsaturated fatty acid:MUFA)は,ラジカルによる傷害を受けにくい.このため,PUFAの増加は細胞のフェロトーシス感受性を高め,MUFAの蓄積はフェロトーシス耐性をもたらす9)(松岡・杉浦の稿).フェロトーシスの誘因となる脂質過酸化は,このように遊離鉄やPUFAで促進される一方,細胞の適切なレドックス制御は過酸化脂質を除去し,フェロトーシスを抑制する10)(酒井らの稿).そのため,フェロトーシスは鉄代謝,脂質代謝,レドックス制御のバランス不均衡がもたらす細胞死と捉えることができる11).
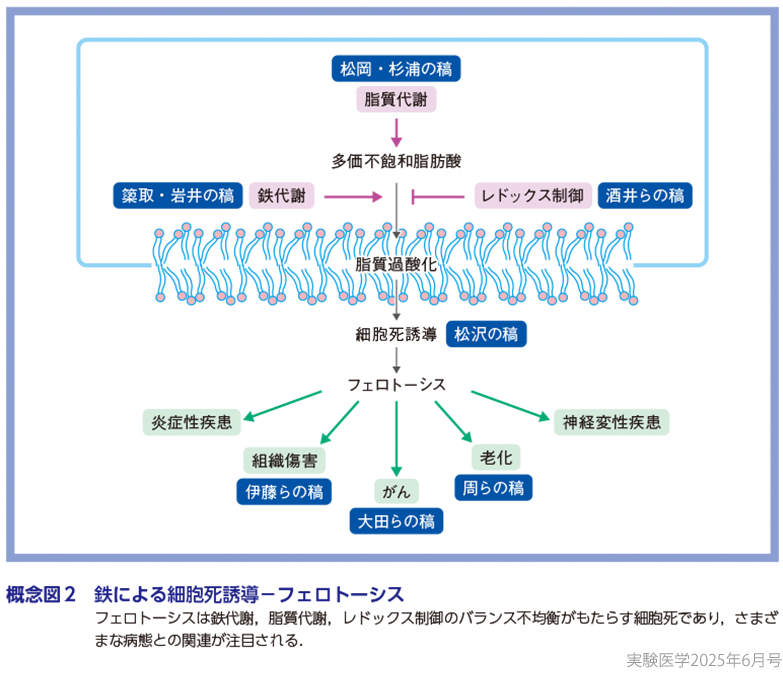
フェロトーシスが脂質過酸化反応を伴う現象であることは明確であるが,過酸化脂質の過剰な蓄積と細胞死の誘導をつなぐ分子実態は十分に明らかになっていない.フェロトーシスはその進行に伴い,細胞膜が崩壊し炎症誘導性の細胞内物質が放出されることが知られている.この過程は一見無秩序で受動的なものに見えるが,近年,細胞膜での過酸化脂質の蓄積に伴って細胞が膨張し,細胞膜の張力が増加することで,機械刺激受容体を介した著しいカチオン流出入が起こることが明らかになった(松沢の稿).このような応答機構の存在は,フェロトーシスが高度に制御された細胞死メカニズムであることを示唆しており,さまざまな生命現象に対するフェロトーシスの積極的な関与が容易に推察される.実際,フェロトーシス研究の拡大により,フェロトーシスが関与するさまざまな生命現象が明らかになってきた12).例えば,フェロトーシスは炎症性疾患や組織傷害(伊藤らの稿),がん(大田らの稿),老化(周らの稿),神経変性疾患などの病態にかかわることが次々に明らかにされ,これらの疾患の病態理解や新規予防・治療法開発における創薬標的としてのフェロトーシスが注目されている13).また,フェロトーシスが生理的な現象であるかは長らくの謎であったが,近年,鳥類の発生過程における筋組織のリモデリングにおいて,フェロトーシスが筋細胞の効率的な除去にかかわることが報告された14).われわれヒトを含む哺乳類においても,発生や成長,組織恒常性維持などのコンテキストにおいて生理的なフェロトーシスが存在するのか,今後の研究が期待される.
一方,このように急速に拡大する研究領域において注意が必要な点として,適切でない実験系の選択やデータの拡大解釈がもたらす概念的な誤解があげられる.現在の定義では,フェロトーシスは鉄のキレート剤および脂質ラジカル捕獲性の抗酸化剤(ferrostatin 1やvitamin Eなど)で抑制される細胞死とされる.また,これらに加えて過酸化脂質やアルデヒドの検出も重要となるであろう15).そのため,フェロトーシスが関与すると考えられる生命現象については,1種類の阻害剤を用いた検証実験のみではなく,これら多面的な視点からの厳格な評価が必要とされる.一方,臨床検体サンプルなど,阻害剤を用いた検証が困難な場合も多く,アポトーシスにおける切断型カスパーゼ3の検出のように,生体内においてフェロトーシスを検出する特異的なバイオマーカーの同定が待たれる.
おわりに
近年,新たな細胞死の概念としてフェロトーシスが発見され,鉄毒性が引き起こす細胞傷害の分子基盤が明確になった.フェロトーシスは鉄代謝のみならず,脂質代謝やレドックス制御とも密接にかかわり,がん,老化,組織傷害などの多岐にわたる研究領域で注目を集めている.それにより,フェロトーシス研究はこれらの研究領域を巻き込みながら拡大しており,これまで個々に研究されてきたこれらの領域がフェロトーシスをキーワードに集約されることで,新たな研究の化学反応が生まれている.今後もこの流れは継続すると考えられ,フェロトーシス研究はより包括的な学問体系へと発展していくと予想される.さらに,鉄は幅広い酵素の分子機能を調節することから,その細胞内動態はほぼすべての細胞機能を制御している可能性があると言っても過言ではない.そのため,フェロトーシス研究の発展にみられるように,鉄が制御する他の生命現象に関する研究も今後大きな発展が期待される.これまで,鉄は主に酵素の補因子として補助的な役割を果たすものと考えられていたが,近年,鉄が能動的に細胞機能を制御する例も報告され,鉄による細胞機能制御の原理の一端が明らかになりつつある.このように,鉄代謝研究は他の分野と相互作用しながら今後ますます拡大し,鉄が関与する新たな生理的・病理的現象の解明につながると期待される.鉄代謝の制御が健康の維持や疾患治療において重要な役割を果たすことが明らかになれば,鉄をターゲットにした新しい治療法や診断法の開発にも貢献するだろう.フェロトーシスをはじめとする鉄代謝の研究は,生命科学の多くの領域での知見を結びつけ,今後の医学やバイオテクノロジーにおける革新的な進展をもたらすと期待される.
鉄研究に魅せられて
筆者が鉄研究と初めて出会ったのは,大学院生として研究の世界に飛び込んでまもなくのことである.当時,FBXL5という機能未知のユビキチンリガーゼについて調べていたところ,この分子が鉄と直接結合する細胞内鉄センサーであり,細胞内の鉄量に応じて自身の安定性を調整し,下流の鉄代謝関連分子の発現量を制御することがわかった.FBXL5は,自身のヘムエリスリンドメインに鉄を結合させるが,このヘムエリスリンは海洋無脊椎動物の血色素として酸素を運搬するタンパク質として知られている.進化の過程で,鉄結合タンパク質とユビキチンリガーゼが融合することにより,このような鉄感知・制御システムが誕生したと考えられる.FBXL5を欠損したマウスは,過剰な鉄が蓄積し,胎生期に死亡してしまうが,その基質であるIRP2を追加で欠損したマウスは,正常に出生し成長する.このような鉄感知・制御システムの美しさと,ノックアウトマウスにみられる劇的な表現型への感動は,筆者がその後も鉄研究を続ける大きなモチベーションとなっている.(諸石寿朗)
文献
- Weiss MC, et al:Nat Microbiol, 1:16116, doi:10.1038/nmicrobiol.2016.116(2016)
- Andreini C, et al:Metallomics, 10:1223-1231, doi:10.1039/c8mt00146d(2018)
- Teh MR, et al:Trends Endocrinol Metab, 35:1026-1049, doi:10.1016/j.tem.2024.04.015(2024)
- Shapiro JS, et al:Nat Cell Biol, 25:1478-1494, doi:10.1038/s41556-023-01225-6(2023)
- Brissot P, et al:Nat Rev Dis Primers, 4:18016, doi:10.1038/nrdp.2018.16(2018)
- Dixon SJ, et al:Cell, 149:1060-1072, doi:10.1016/j.cell.2012.03.042(2012)
- Stockwell BR, et al:Cell, 171:273-285, doi:10.1016/j.cell.2017.09.021(2017)
- Galy B, et al:Nat Rev Mol Cell Biol, 25:133-155, doi:10.1038/s41580-023-00648-1(2024)
- Pope LE & Dixon SJ:Trends Cell Biol, 33:1077-1087, doi:10.1016/j.tcb.2023.05.003(2023)
- Halliwell B:Nat Rev Mol Cell Biol, 25:13-33, doi:10.1038/s41580-023-00645-4(2024)
- Stockwell BR:Cell, 185:2401-2421, doi:10.1016/j.cell.2022.06.003(2022)
- Jiang X, et al:Nat Rev Mol Cell Biol, 22:266-282, doi:10.1038/s41580-020-00324-8(2021)
- Sun S, et al:Signal Transduct Target Ther, 8:372, doi:10.1038/s41392-023-01606-1(2023)
- Co HKC, et al:Nature, 631:654-662, doi:10.1038/s41586-024-07623-6(2024)
- Chen Z, et al:Heliyon, 10:e23507, doi:10.1016/j.heliyon.2023.e23507(2024)
本記事のDOI:10.18958/7737-00001-0006024-00
著者プロフィール
諸石寿朗:2008年九州大学医学部卒業.九州大学大学院医学系学府を経て,’13〜’17年に米国カリフォルニア大学サンディエゴ校に留学.帰国後,’17年熊本大学 准教授,’20年熊本大学 教授を経て,’24年より現職の東京科学大学 教授.専門は分子生物学・細胞生物学.多細胞生物において細胞の多様性はどのように生まれ,どう変容するのか,また,そこにどのような意義があるのか? 多種多様な細胞から構成される組織の恒常性を維持するしくみを理解したいと考えており,サイエンスの面白さをともに追求していける仲間を募集しています.趣味は水辺のいきもの探索とアクアリウム.










